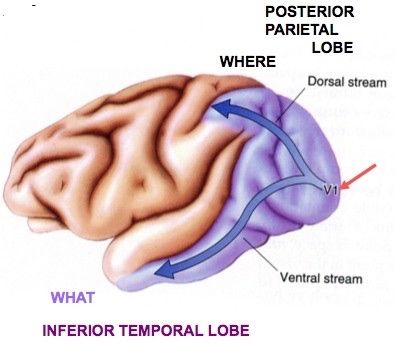
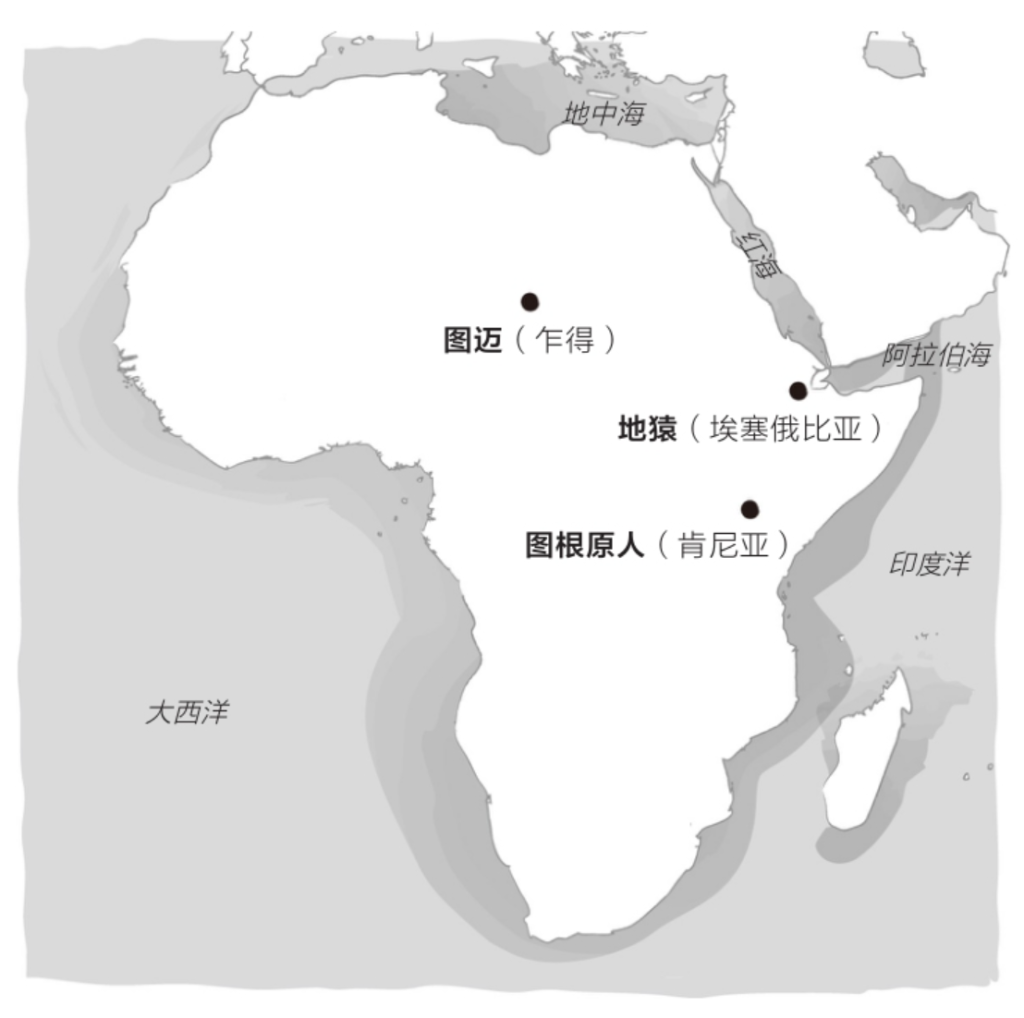
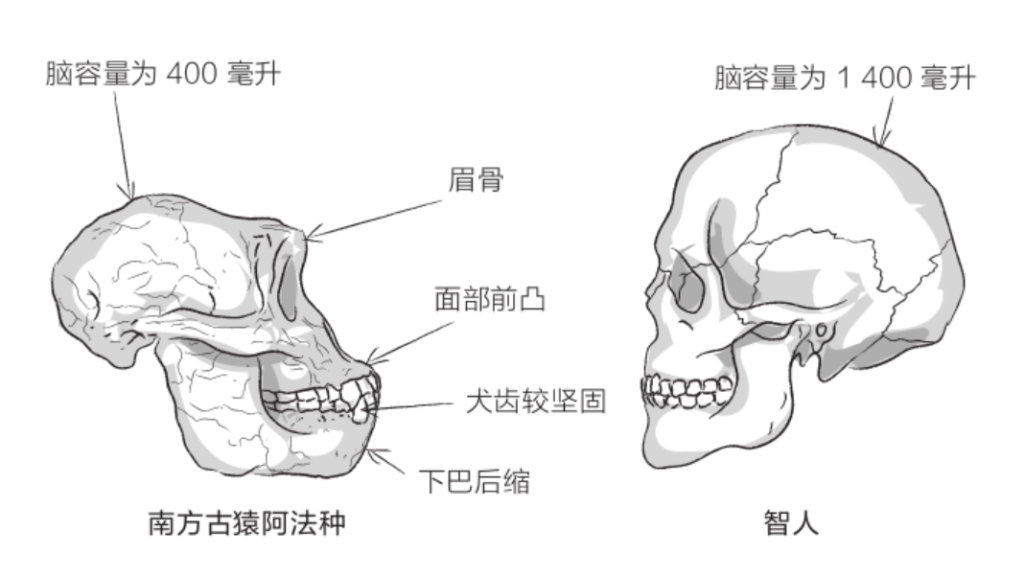
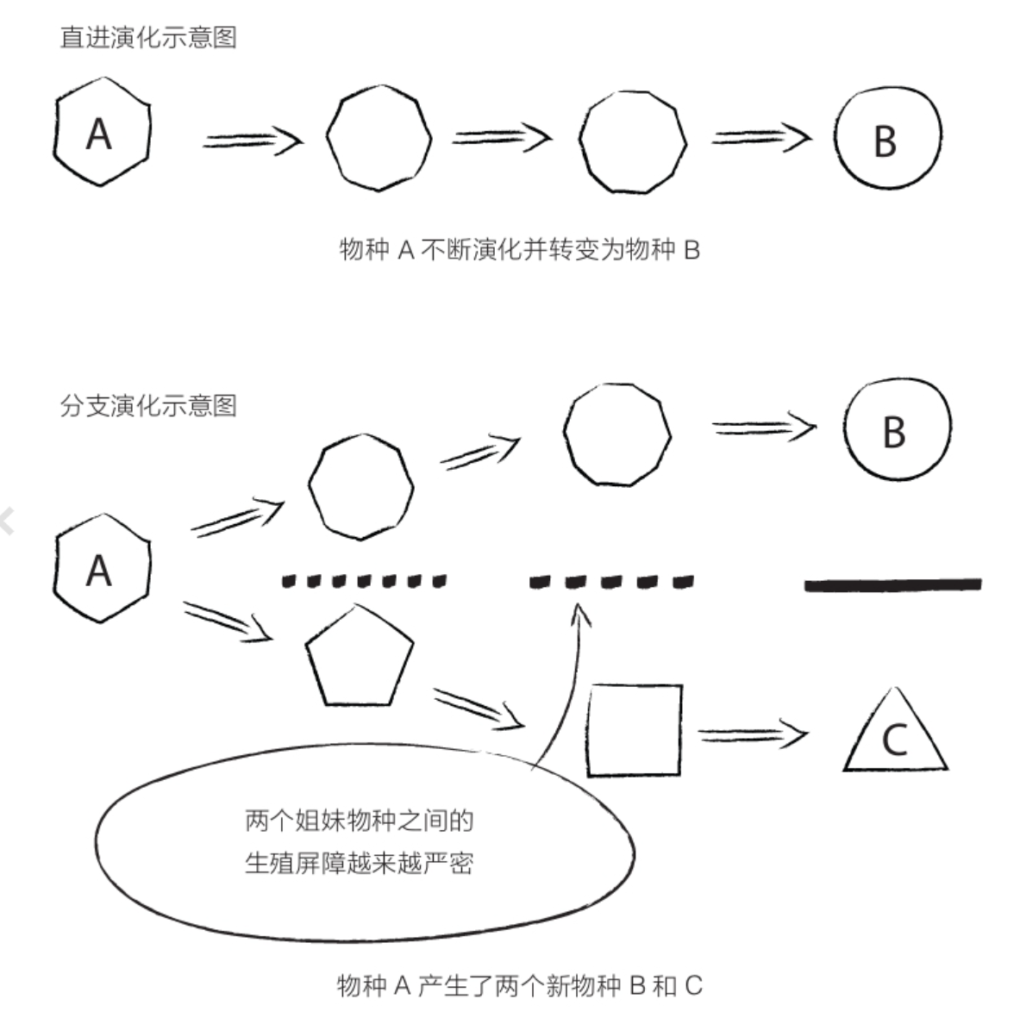
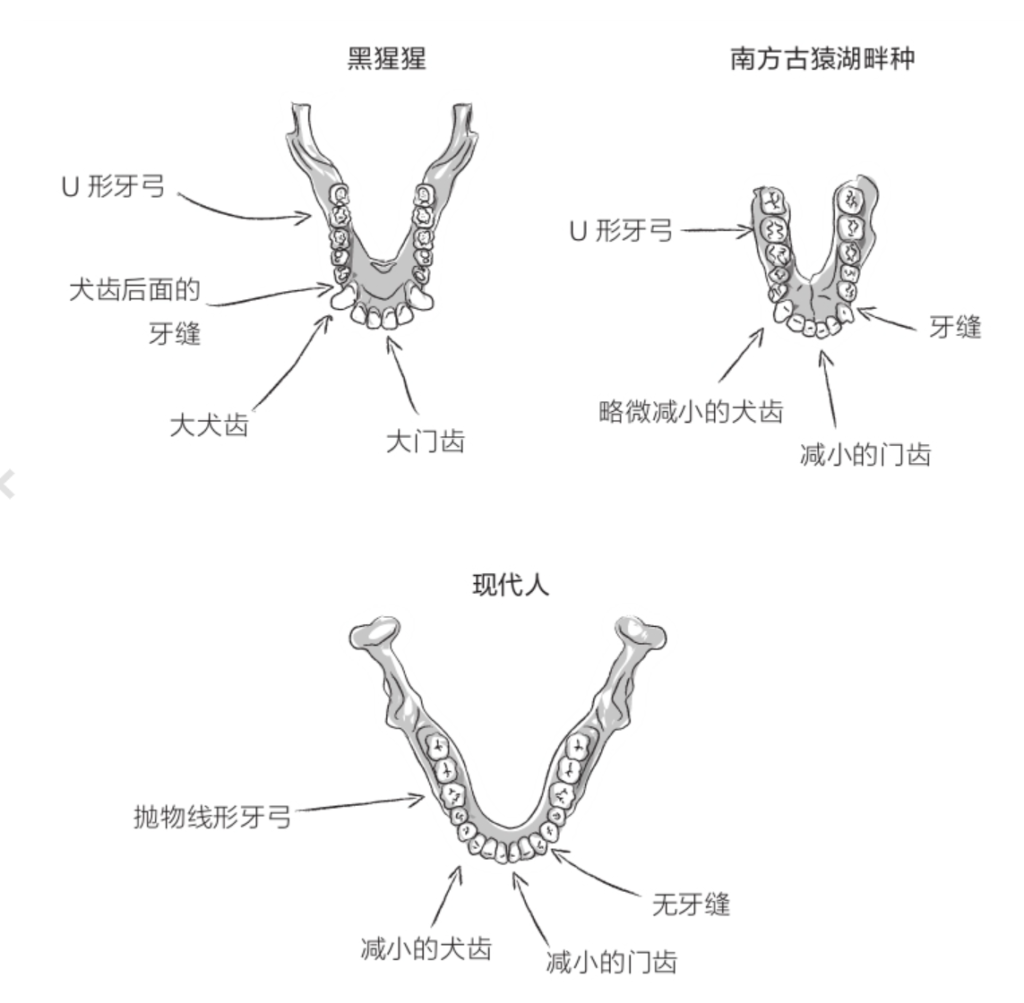
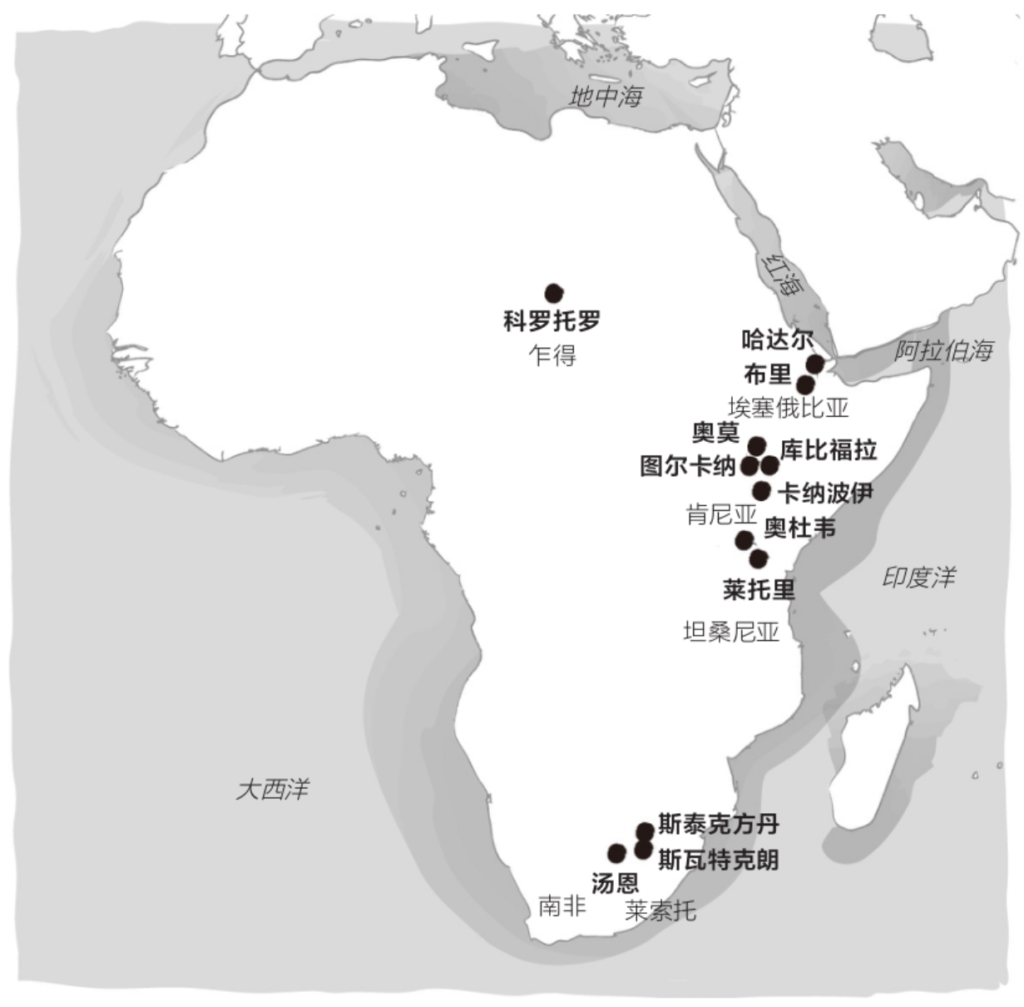
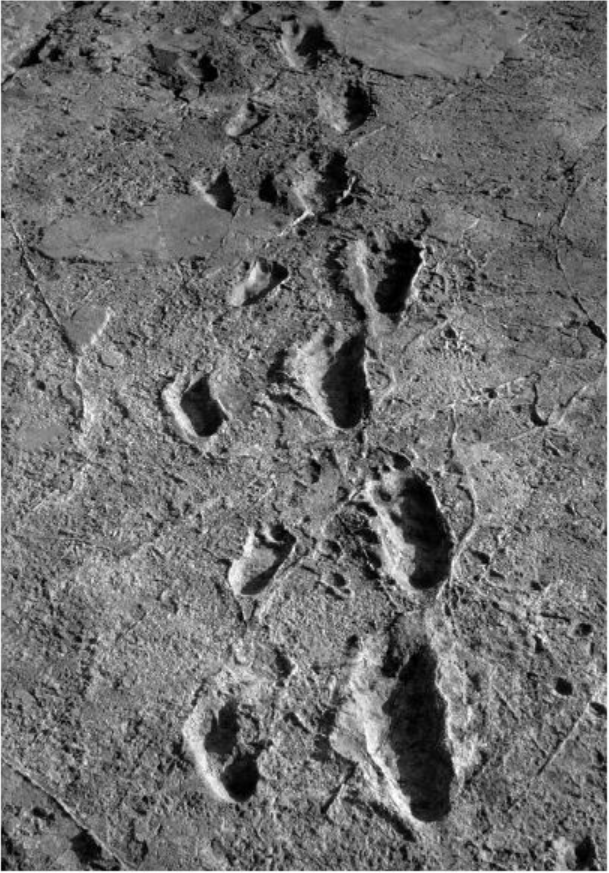
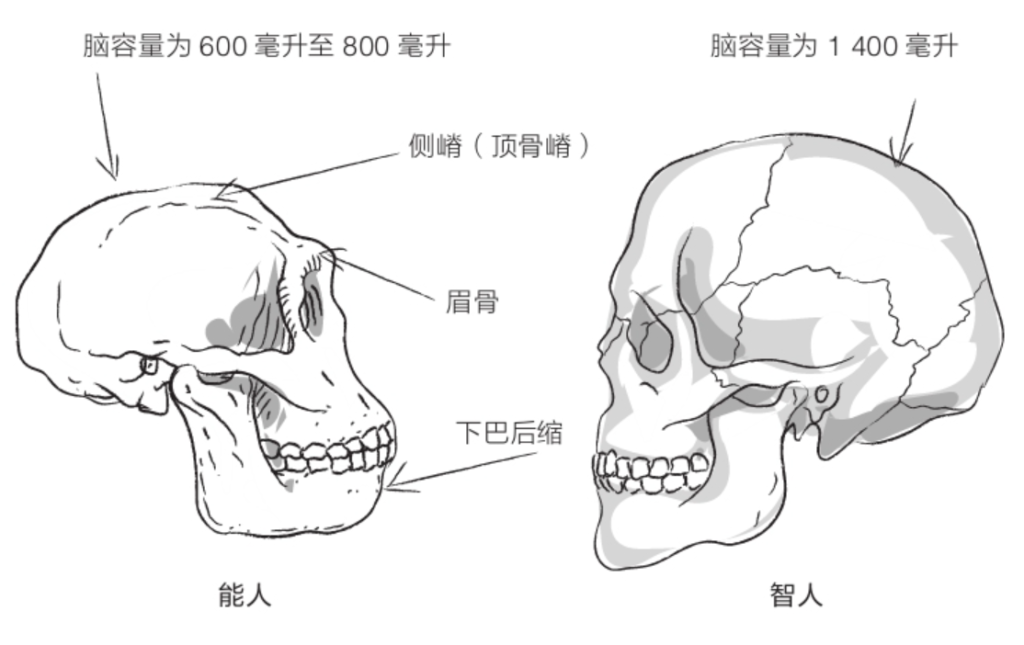
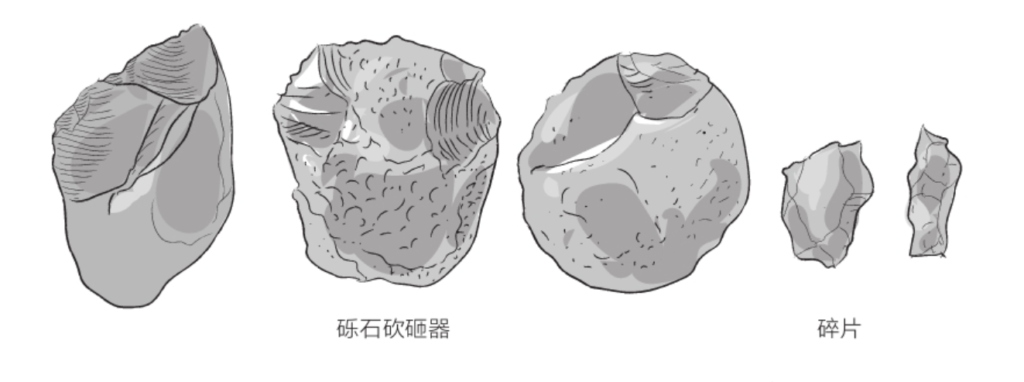
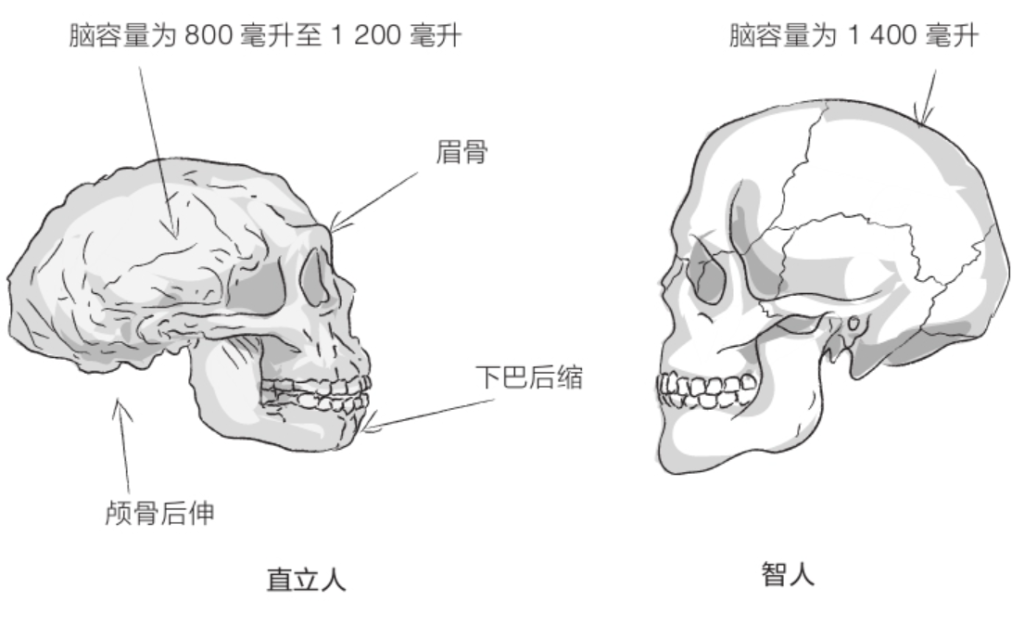
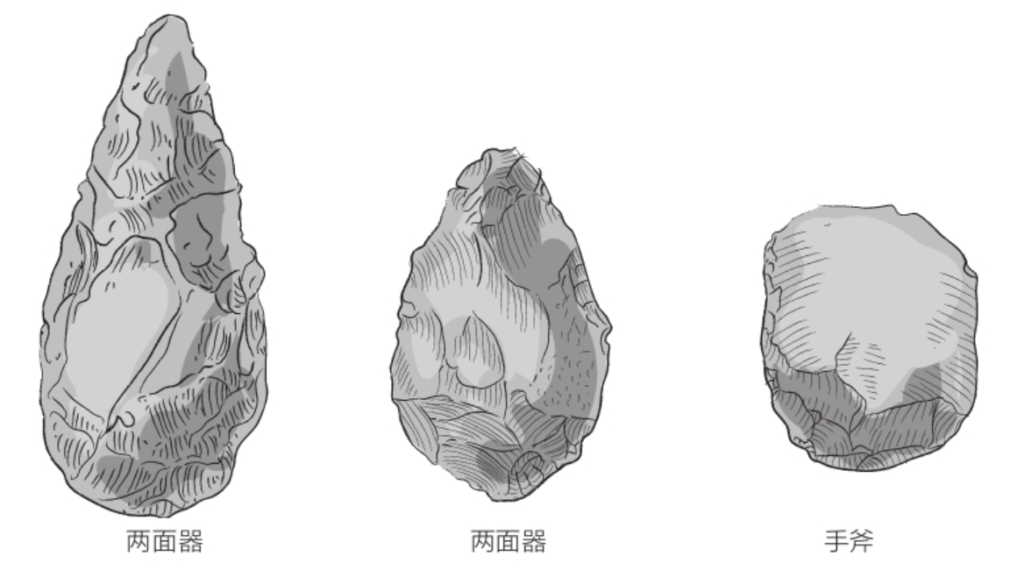
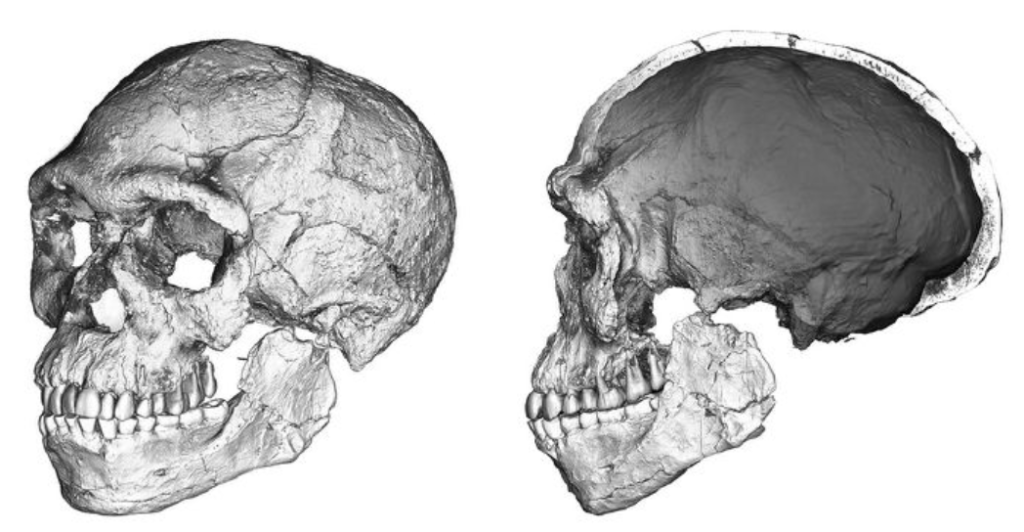
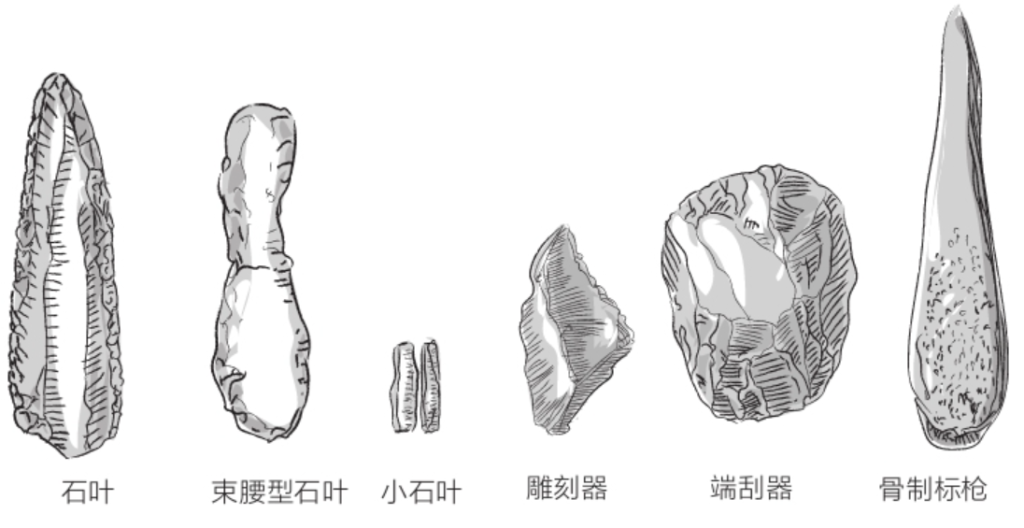
自由心证原则之确立以人类理性觉醒与确定性认知为前提,在法官内心确信中体现为对心证盖然性的承认和盖然性程度确证的理解差异。1927年,帝国法院首次在RGSt 61, 202判决中确立了刑事案件定罪意义上的内心确信,并提出以高度盖然性(die hohe Wahrscheinlichkeit)作为自由心证内心确信的标准。此种观点的核心要旨在于,在纯粹主观主义的自由心证原则之中引入客观主义底色的概率判断,即高度盖然性,与此同时强调不通过主观因素过分夸大“高度盖然性”,而是通过推定降低真相所要求的盖然性程度。虽然判决中并未涉及证明标准的维度,但至少明确确立了证明标准是基于诉讼材料所获得的高度盖然性,此种高度盖然性由法官基于理性推理而来,代表着自由心证的客观化。
CONTENTS 1 Introduction 2 GPT Diffusion Theory 3 The First Industrial Revolution and Britain’s Rise 4 The Second Industrial Revolution and America’s Ascent 5 Japan’s Challenge in the Third Industrial Revolution 6 A Statistical Analysis of Software Engineering Skill Infrastructure and Computerization 7 US-China Competition in AI and the Fourth Industrial Revolution 8 Conclusion
1 Introduction
IN JULY 2018, the BRICS nations (Brazil, Russia, India, China, and South Africa) convened in Johannesburg around a specific, noteworthy theme: “Collaboration for Inclusive Growth and Shared Prosperity in the Fourth Industrial Revolution.” The theme was noteworthy in part because of its specificity. Previous iterations of the BRICS summit, which gathers five nations that account for about 40 percent of the world’s population and 25 percent of the world’s GDP,1 had tackled fuzzy slogans such as “Stronger Partnership for a Brighter Future” and “Broad Vision, Shared Prosperity.” What stood out not only about that year’s theme but also in comments by BRICS leaders at the summit was an unambiguous conviction that the world was undergoing a momentous season of technological change—one warranting the title “Fourth Industrial Revolution.”2
Throughout the gathering, leaders of these five major emerging economies declared that the ongoing technological transition represented a rare opportunity for accelerating economic growth. When Chinese president Xi Jinping addressed the four other leaders of major emerging economies, he laid out the historical stakes of that belief:
From the mechanization of the first industrial revolution in the 18th century, to the electrification of the second industrial revolution in the 19th century, to the informatization of the third industrial revolution in the 20th century, rounds of disruptive technological innovation have … fundamentally changed the development trajectory of human history.3
Citing recent breakthroughs in cutting-edge technologies like artificial intelligence (AI), Xi proclaimed, “Today, we are experiencing a larger and deeper round of technological revolution and industrial transformation.”4
While the BRICS summit did not explicitly address how the Fourth Industrial Revolution could reshape the international economic order, the implications of Xi’s remarks loomed in the backdrop. In the following months, Chinese analysts and scholars expanded upon them, especially the connection he drew between technological disruption and global leadership transitions.5 One commentary on Xi’s speech, published on the website of the authoritative Chinese Communist Party publication Study Times, detailed the geopolitical consequences of past technological revolutions: “Britain seized the opportunity of the first industrial revolution and established a world-leading productivity advantage.… After the second industrial revolution, the United States seized the dominance of advanced productivity from Britain.”6 In his analysis of Xi’s address, Professor Jin Canrong of Renmin University, an influential Chinese international relations scholar, argued that China has a better chance than the United States of winning the competition over the Fourth Industrial Revolution.7
This broad sketch of power transition by way of technological revolution also resonates with US policymakers and leading thinkers. In his first press conference after taking office, President Joe Biden underscored the need to “own the future” as it relates to competition in emerging technologies, pledging that China’s goal to become “the most powerful country in the world” was “not going to happen on [his] watch.”8 In 2018, the US Congress stood up the National Security Commission on Artificial Intelligence (NSCAI), an influential body that convened leading government officials, technology experts, and social scientists to study the national security implications of AI. Comparing AI’s possible impact to past technologies like electricity, the NSCAI’s 756-page final report warned that the United States would soon lose its technological leadership to China if it did not adequately prepare for the “AI revolution.”9
Caught up in the latest technical advances coming out of Silicon Valley or Beijing’s Zhongguancun, these sweeping narratives disregard the process by which emerging technologies can influence a power transition. How do technological revolutions affect the rise and fall of great powers? Is there a discernible pattern that characterizes how previous industrial revolutions shaped the global balance of power? If such a pattern exists, how would it inform our understanding of the Fourth Industrial Revolution and US-China technological competition?
Conventional Wisdom on Technology-Driven Power Transitions
International relations scholars have long observed the link between disruptive technological breakthroughs and the rise and fall of great powers.10 At a general level, as Yale historian Paul Kennedy has established, this process involves “differentials in growth rates and technological change, leading to shifts in the global economic balances, which in turn gradually impinge upon the political and military balances.”11 Yet, as is the case with present-day speculation about the effects of new technologies on the US-China power balance, largely missing from the international relations literature is an explanation of how technological change creates the conditions for a great power to leapfrog its rival. Scholars have carefully scrutinized how shifts in economic balances affect global military power and political leadership, but there is a need for further investigation into the very first step of Kennedy’s causal chain: the link between technological change and differentials in long-term growth rates among great powers.12
Among studies that do examine the mechanics of how technological change shapes economic power transitions, the standard explanation stresses dominance over critical technological innovations in new, fast-growing industries (“leading sectors”). Britain became the world’s most productive economy, according to this logic, because it was home to new advances that transformed its burgeoning textile industry, such as James Hargreaves’s spinning jenny. In the same vein, Germany’s mastery of major breakthroughs in the chemical industry is seen as pivotal to its subsequent challenge to British economic leadership. Informed by historical analysis, the leading-sector (LS) perspective posits that, during major technological shifts, the global balance of economic power tips toward “the states which were the first to introduce the most important innovations.”13
Why do the benefits of leading sectors accrue to certain countries? Explanations vary, but most stress the goodness-of-fit between a nation’s domestic institutions and the demands of disruptive technologies. At a general level, some scholars argue that rising powers quickly adapt to new leading sectors because they are unburdened by the vested interests that have built up in more established powers.14 Others point to more specific factors, including the degree of government centralization or sectoral governance arrangements.15 Common to all these perspectives is a focus on the institutions that allow one country to first introduce major breakthroughs in an emerging industry. In the case of Britain’s rise, for example, many influential histories highlight institutions that supported “heroic” inventors.16 Likewise, accounts of Germany’s success with leading sectors focus on its investments in scientific education and industrial research laboratories.17
The broad outlines of LS theory exert substantial influence in academic and policymaking circles. Field-defining texts, including works by Robert Gilpin and Paul Kennedy, use the LS model to map out the rise and fall of great powers.18 In a review of international relations scholarship, Daniel Drezner summarizes their conclusions: “Historically, a great power has acquired hegemon status through a near-monopoly on innovation in leading sectors.”19
The LS template also informs contemporary discussion of China’s challenge to US technological leadership. In another speech about how China could leverage this new round of industrial revolution to become a “science and technology superpower,” President Xi called for China to develop into “the world’s primary center for science and high ground for innovation.”20 As US policymakers confront China’s growing strength in emerging technologies like AI, they also frame the competition in terms of which country will be able to generate radical advances in new leading sectors.21
Who did it first? Which country innovated it first? Presented with technical breakthroughs that inspire astonishment, it is only natural to gravitate toward the moment of initial discovery. When today’s leaders evoke past industrial revolutions, as Xi did in his speech to the BRICS nations, they tap into historical accounts of technological progress that also center the moment of innovation.22 The economist and historian Nathan Rosenberg diagnoses the problem with these innovation-centric perspectives: “Much less attention … if any at all, has been accorded to the rate at which new technologies have been adopted and embedded in the productive process. Indeed the diffusion process has often been assumed out of existence.”23 Yet, without the humble undertaking of diffusion, even the most extraordinary advances will not matter.
Taking diffusion seriously leads to a different explanation for how technological revolutions affect the rise and fall of great powers. A diffusion-centric framework probes what comes after the hype. Less concerned with which state first introduced major innovations, it instead asks why some states were more successful at adapting and embracing new technologies at scale. As outlined in the next section, this alternative pathway points toward a different set of institutional factors that underpin leadership in times of technological leadership, in particular institutions that widen the base of engineering skills and knowledge linked to foundational technologies.
GPT Diffusion Theory
In September 2020, the Guardian published an opinion piece arguing that humans should not fear new breakthroughs in AI. Noting that “Stephen Hawking has warned that AI could ‘spell the end of the human race,’ ” the article’s “author” contends that “I am here to convince you not to worry. Artificial intelligence will not destroy humans. Believe me.”24 If one came away from this piece with the feeling that the author had a rose-tinted view of the future of AI, it would be a perfectly reasonable judgment. After all, the author was GPT-3, an AI model that can understand and produce humanlike text.
Released earlier that year by OpenAI, a San Francisco–based AI lab, GPT-3 surprised everyone—including its designers—with its versatility. In addition to generating poetry and essays like the Guardian op-ed from scratch, early users demonstrated GPT-3’s impressive capabilities in writing code, translating languages, and building chatbots.25 Six months after its launch, one compilation listed sixty-six unique use cases of GPT-3, which ranged from automatically updating spreadsheets to generating website landing pages.26 Two years later, OpenAI’s acclaimed ChatGPT model, built on an improved version of GPT-3, would set the internet aflame with its wide-ranging capabilities.27
While the name “GPT-3” derives from a class of language models known as “generative pre-trained transformers,” the abbreviation, coincidentally, also speaks to the broader significance of recent breakthroughs in AI: the possible arrival of the next general-purpose technology (GPT). Foundational breakthroughs in the ability of computers to perform tasks that usually require human intelligence have the potential to transform countless industries. Hence, scholars and policymakers often compare advances in AI to electricity, the prototypical GPT.28 As Kevin Kelly, the former editor of WIRED, once put it, “Everything that we formerly electrified we will now cognitize … business plans of the next 10,000 startups are easy to forecast: Take X and add AI.”29
In this book, I argue that patterns in how GPTs diffuse throughout the economy illuminate a novel explanation for how and when technological changes affect power transitions. The emergence of GPTs—fundamental advances that can transform many application sectors—provides an opening for major shifts in economic leadership. Characterized by their scope for continuous improvement, pervasive applicability across the economy, and synergies with other technological advances, GPTs carry an immense potential for boosting productivity.30 Carefully tracking how the various applications of GPTs are adopted across various industries, a process I refer to as “GPT diffusion,” is essential to understanding how technological revolutions disrupt economic power balances.
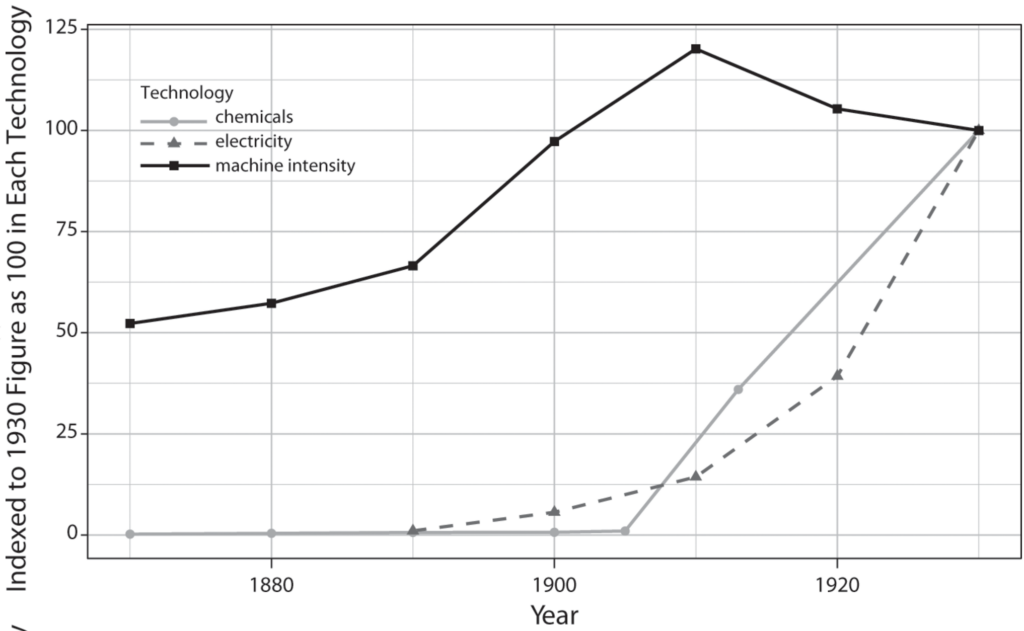
Based on the experience of past GPTs, this potential productivity boost comes with one notable caveat: the full impact of a GPT manifests only after a gradual process of diffusion into pervasive use.31 GPTs demand structural changes across a range of technology systems, which involve complementary innovations, organizational adaptations, and workforce adjustments.32 For example, electrification’s boost to productivity materialized about five decades after the introduction of the first electric dynamo, occurring only after factories had restructured their layouts and there had been interrelated breakthroughs in steam turbines.33 Fittingly, after the release of GPT-3, OpenAI CEO Sam Altman alluded to this extended trajectory: “The GPT-3 hype is way too much … it still has serious weaknesses and sometimes makes very silly mistakes. AI is going to change the world, but GPT-3 is just a very early glimpse. We have a lot still to figure out.”34
Informed by historical patterns of GPT diffusion, my explanation for technology-driven power transitions diverges significantly from the standard LS account. Specifically, these two causal mechanisms differ along three key dimensions, which relate to the technological revolution’s impact timeframe, phase of relative advantage, and breadth of growth. First, while the GPT mechanism involves a protracted gestation period between a GPT’s emergence and resulting productivity boosts, the LS mechanism assumes that there is only a brief window during which countries can capture profits in leading sectors. “The greatest marginal stimulation to growth may therefore come early in the sector’s development at the time when the sector itself is expanding rapidly,” William Thompson reasons.35 By contrast, the most pronounced effects on growth arrive late in a GPT’s development.
Second, the GPT and LS mechanisms also assign disparate weights to innovation and diffusion. Technological change involves a phase when the technology is first incubated as a viable commercial application (“innovation”) and a phase when the innovation permeates across a population of potential users (“diffusion”). The LS mechanism is primarily concerned about which country dominates innovation in leading sectors, capturing the accompanying monopoly profits.36 Under the GPT mechanism, successful adaptation to technological revolutions is less about being the first to introduce major innovations and more about effectively adopting GPTs across a wide range of economic sectors.
Third, regarding the breadth of technological transformation and economic growth, the LS mechanism focuses on the contributions of a limited number of leading sectors and new industries to economic growth in a particular period.37 In contrast, GPT-fueled productivity growth is spread across a broad range of industries.38 Dispersed productivity increases from many industries and sectors come from the extension and generalization of localized advances in GPTs.39 Thus, the LS mechanism expects the breadth of growth in a particular period to be concentrated in leading sectors, whereas the GPT mechanism expects technological complementarities to be dispersed across many sectors.
A clearer understanding of the contours of technological change in times of economic power transition informs which institutional variables matter most. If the LS trajectory holds, then the most important institutional endowments and responses are those that support a monopoly on innovation in leading sectors. In the context of skill formation, institutional competencies in science and basic research gain priority. For instance, the conventional explanation of Germany’s industrial rise in the late nineteenth century attributes its technological leadership to investments in industrial research labs and highly skilled chemists. These supported Germany’s dominance of the chemical industry, a key LS of the period.40
The impact pathway of GPTs brings another set of institutional complementarities to the fore. GPT diffusion theory highlights the importance of “GPT skill infrastructure”: education and training systems that widen the pool of engineering skills and knowledge linked to a GPT. When widespread adoption of GPTs is the priority, it is ordinary engineers, not heroic inventors, who matter most. Widening the base of engineering skills associated with a GPT cultivates a more interconnected technological system, spurring cross-fertilization between institutions optimized for applied technology and those oriented toward foundational research.41
Returning to the example of late-nineteenth-century advances in chemicals, GPT diffusion spotlights institutional adjustments that differ from those of the LS mechanism. In a decades-long process, innovations in chemical engineering practices gradually enabled the chemicalization of procedures common to many industries beyond synthetic dyes, which was controlled by Germany. Despite trailing Germany in the capacity to produce elite chemists and frontier chemical research, the United States was more effective at adapting to chemicalization because it first institutionalized the discipline of chemical engineering.42
Of course, since GPT diffusion depends on factors aside from human capital, GPT skill infrastructure represents one of many institutional forces at work. Standards-setting organizations, financing bodies, and the competitiveness of markets can all influence the flow of information between the GPT domain and application sectors.43 Since institutions of skill formation produce impacts that spill over into and complement other institutional arrangements, they comprise the focus of my analysis.44
Assessing GPT Diffusion across Industrial Revolutions
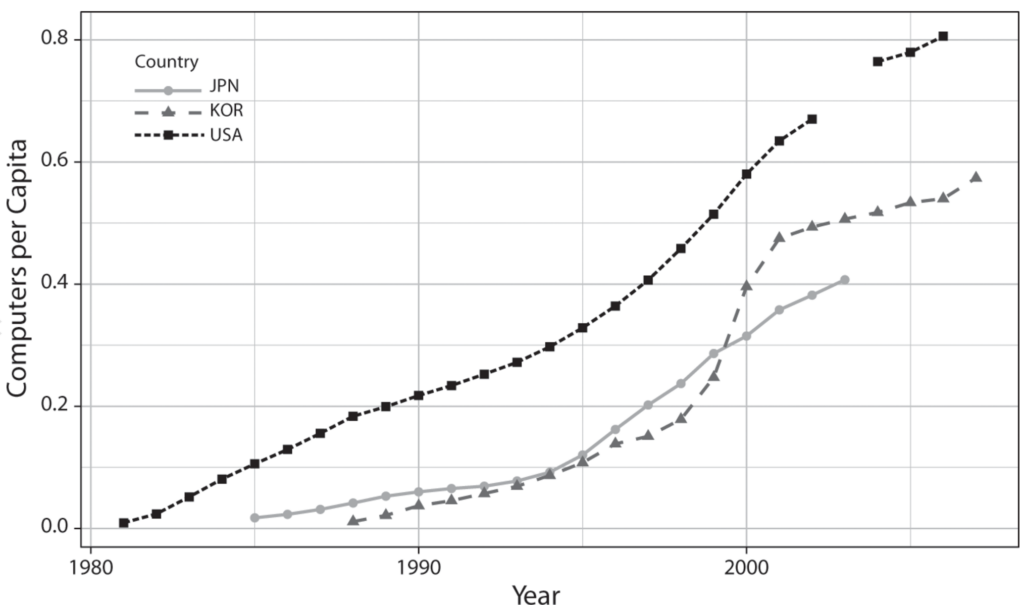
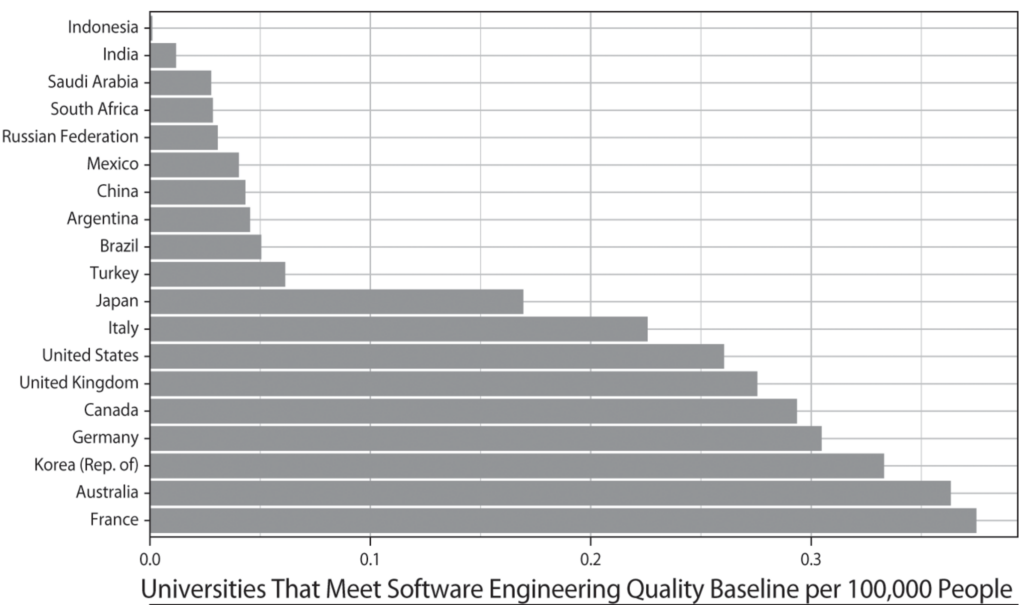
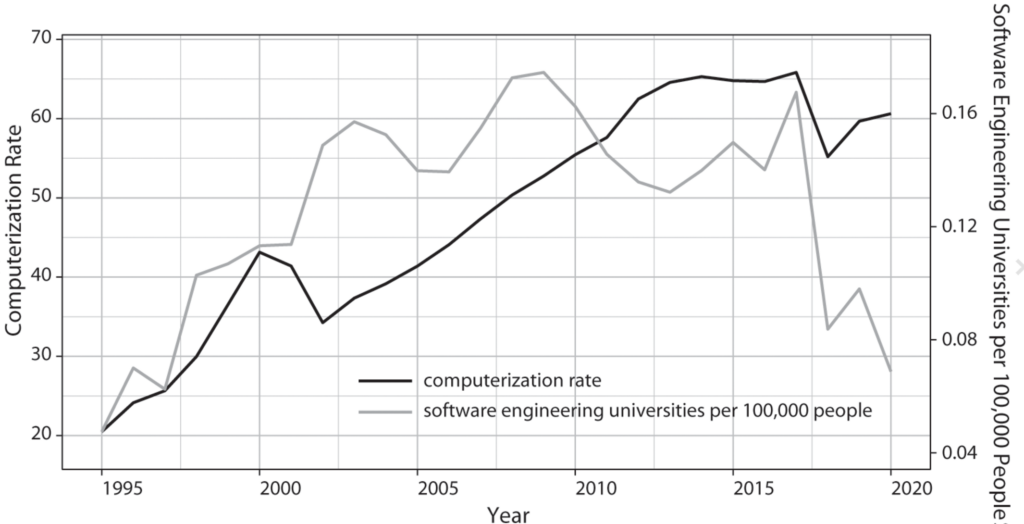
To test this argument, I employ a mixed-methods approach that pairs qualitative historical analysis with quantitative methods. Historical case studies permit me to thoroughly trace the interactions between technologies and institutions among great powers in previous industrial revolutions. I then explore the generalizability of GPT diffusion theory beyond the chosen set of great powers. Using data on nineteen countries from 1995 to 2020, I analyze the theorized connection between GPT skill infrastructure in software engineering and computerization rates.
To investigate the causal processes that connect technological changes to economic power transitions, I set the LS mechanism against the GPT diffusion mechanism across three historical case studies: Britain’s rise to preeminence in the First Industrial Revolution (IR-1); America’s and Germany’s overtaking of Britain in the Second Industrial Revolution (IR-2); and Japan’s challenge to America’s technological dominance in the Third Industrial Revolution (IR-3), or what is sometimes called the “information revolution.” This case setup allows for a fair and decisive assessment of the explanatory relevance of GPT diffusion theory in comparison to LS theory. Because the IR-1 and IR-2 function as typical cases where the cause and outcome are clearly present, they are ideal for developing and testing mechanism-based theories.45 The IR-3, a deviant case in that a technological revolution is not followed by an economic power transition, provides a different but still useful way to compare the two mechanisms.
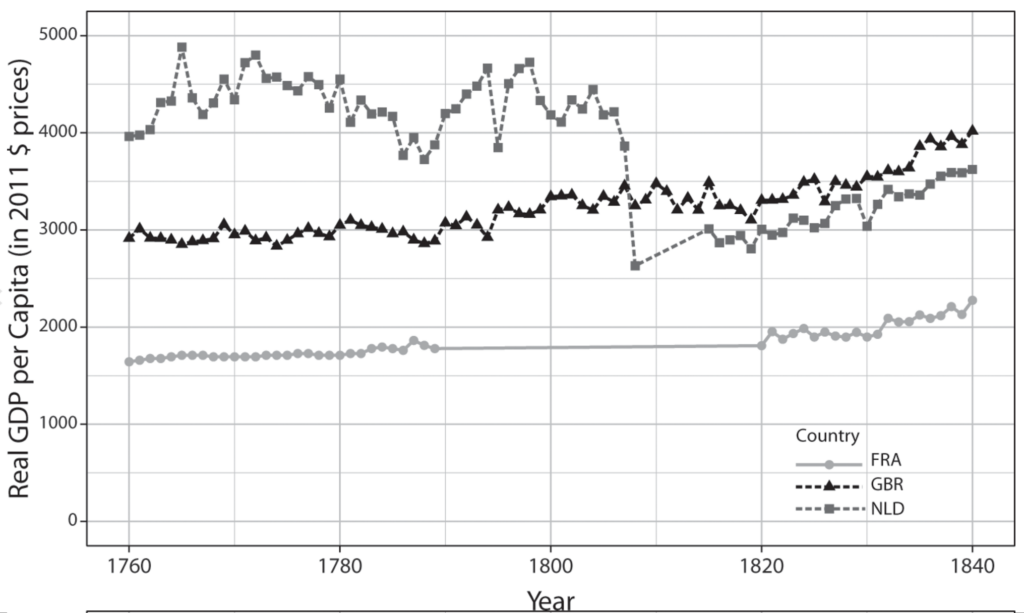
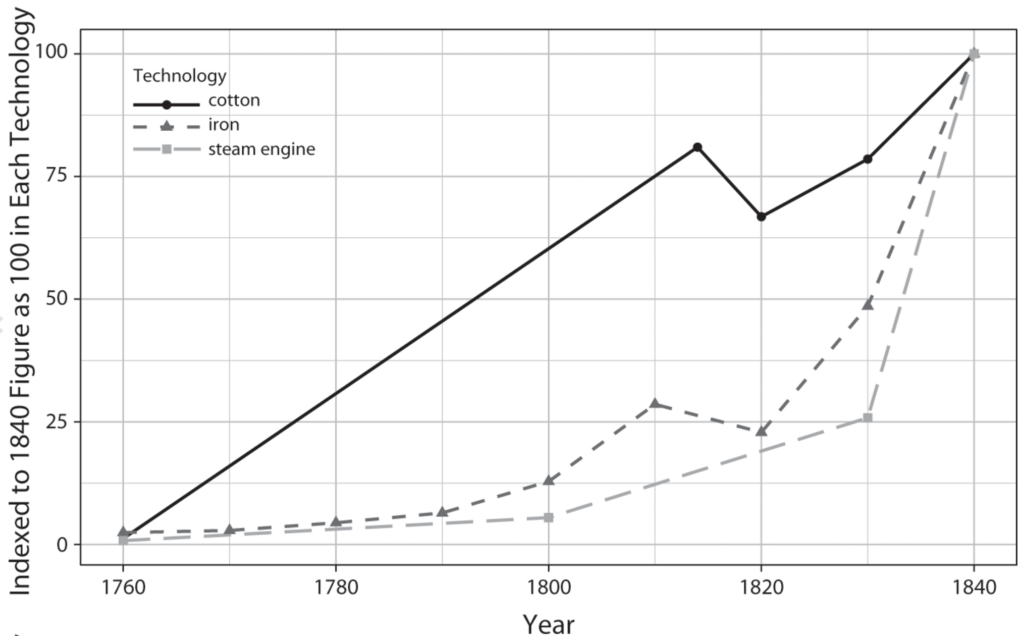
The IR-1 (1780–1840) is a paradigmatic case of technology-driven power transition. It is well established that the IR-1’s technological advances propelled Great Britain to unrivaled economic supremacy. As for the specific causal pathway, international relations scholarship tends to attribute Britain’s rise to its monopoly over innovation in cotton textiles and other leading sectors. According to these accounts, Britain’s technological leadership in the IR-1 sprang from its institutional capacity to nurture genius inventors in these sectors. Since the publication of these field-defining works, economic and technology historians have uncovered that the impacts on British industrialization of the two most prominent areas of technological change, cotton textiles and iron, followed different trajectories. Often relying on formal econometric methods to understand the impact of key technologies, these historical accounts question the prevailing narrative of the IR-1.
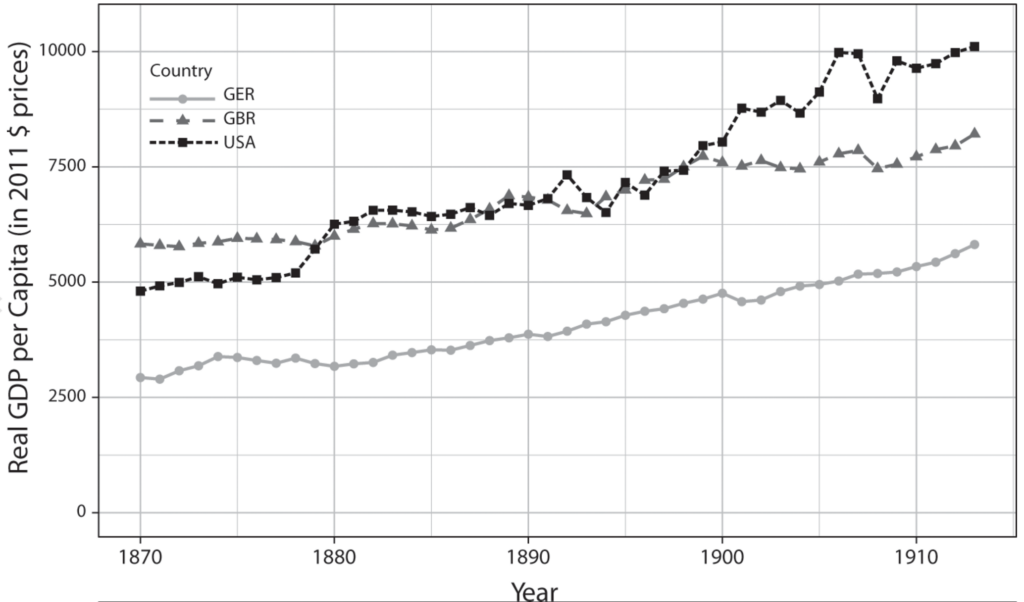
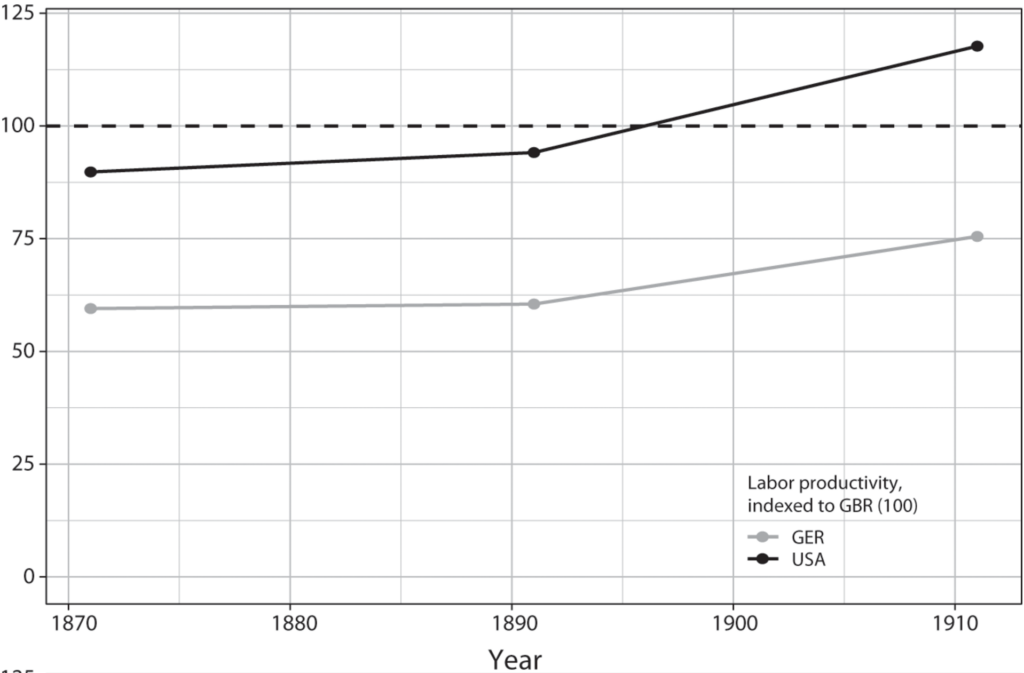
The IR-2 (1870–1914) supplies another opportunity to pit GPT diffusion theory against the LS account. International relations scholars interpret the IR-2 as a case in which Britain’s rivals challenged its economic leadership because they first introduced significant technological advances in leading sectors. Particular emphasis is placed on Germany’s ability to corner market shares in chemicals, which is linked to its strengths in scientific education and industrial research institutions. More granular data on cross-national differences in engineering education suggest that the U.S. technological advantage rested on the country’s wide base of mechanical engineers. Combined with detailed tracing of the pace and extent of technology adoption during this period, this chapter’s evidence suggests modifications to conventional understandings of the IR-2.
In the IR-3 (1960–2000), fundamental breakthroughs in information and communication technologies presented another opening for a shift in economic leadership. During this period, prominent thinkers warned that Japan’s lead in industries experiencing rapid technological change, including semiconductors and consumer electronics, would threaten U.S. economic leadership. Influential scholars and policymakers advocated for the United States to adopt Japan’s keiretsu system of industrial organization and its aggressive industrial policy approach. Ultimately, Japan’s productivity growth stalled in the 1990s. Given the absence of an economic power transition, the primary function of the IR-3 case therefore is to provide disconfirming evidence of the two explanations. If the components of the LS mechanism were present, then the fact that an economic power transition did not occur would damage the credibility of the LS mechanism. The same condition applies to GPT diffusion theory.
In each of the cases, I follow the same standardized procedures. First, I test three pairs of competing propositions about the key technological trajectories, derived from the different expectations of the LS and GPT mechanisms related to the impact timeframe, phase of relative advantage, and breadth of growth. Then, depending on whether the LS or GPT trajectory better accords with the historical evidence, I analyze the goodness-of-fit between the institutional competencies of leading industrial powers and the prevailing trajectory. For instance, if an industrial revolution is better characterized by the GPT trajectory, then the corresponding case analysis should show that differences in GPT skill infrastructure determine which powers rise and fall. Although I primarily distinguish GPT diffusion theory from the LS model, I also examine alternative factors unique to the particular case, as well as two other prominent explanations of how advanced economies differentially benefit from technological changes (the varieties of capitalism and threat-based approaches).
The historical case analysis supports the explanatory power of the GPT mechanism over the LS mechanism. In all three periods, technological changes affected the rise and fall of great powers in a gradual, decades-long impact pathway that advantaged those that effectively diffused GPTs across a broad range of sectors. Education and training systems that cultivated broad pools of engineering skills proved crucial to GPT diffusion.
Evaluating these two competing explanations requires a clear understanding of the cause and outcome that bracket both the GPT and LS mechanisms. The hypothesized cause is a “technological revolution,” or a period characterized by particularly disruptive technological advances.46 Since the shape of technological change is uneven, not all improvements in useful knowledge are relevant for power transitions.47 However, some extraordinary clusters of technological breakthroughs, often deemed industrial revolutions by historians, do have ramifications for the rise and fall of great powers.48 I am primarily interested in the pathway by which these technological revolutions influence the global distribution of power.
The outcome variable of interest is an economic power transition, in which one great power sustains productivity growth at higher levels than its rivals. The balance of power can shift in many ways; here I focus on relative economic growth rates because they are catalysts for intensifying hegemonic rivalries.49 Productivity growth, in particular, determines economic growth over the long run. Unique in its fungibility with other forms of power, sustained economic growth is central to a state’s ability to exert political and military influence. As demonstrated by the outcomes of interstate conflicts between great powers, economic and productive capacity is the foundation of military power.50
Lastly, the quantitative analysis supplements the historical case studies by scrutinizing the generalizability of GPT diffusion theory outside of great powers. A key observable implication of my argument is that the rate at which a GPT spreads throughout the economy owes much to that country’s institutional capacity to widen the pool of pertinent engineering skills and knowledge. Using a novel method to estimate the breadth of software engineering education at a cross-national level, I analyze the theorized connection between GPT skill infrastructure and computerization rates across nineteen advanced and emerging economies from 1995 to 2020. I supplement my time-series cross-sectional models with a duration analysis and cross-sectional regressions. Robust to many alternative specifications, my results show that, at least for computing technology, advanced economies that have higher levels of GPT skill infrastructure preside over higher rates of GPT diffusion.
Key Contributions
The book makes several contributions to scholarship on power transitions and the effects of technological change on international politics. First, it puts forward a novel explanation for how and when significant technological breakthroughs generate a power transition in the international system. GPT diffusion theory revises the dominant theory based on leading sectors, which holds significant sway over academic and policymaking circles. By deepening our understanding of how technological revolutions influence shifts in economic leadership, this book also contributes to long-standing debates about the causes of power transitions.51
Second, the findings of this book bear directly on present-day technological competition between the United States and China. Emphasizing where fundamental breakthroughs are first seeded, the LS template strongly informs not only assessments of the US-China competition for technological leadership but also the ways in which leading policymakers in both countries formulate technology strategies. It is no coincidence that the three cases in this study match the three technological revolutions referenced by Chinese president Xi in his speech on the IR-4 to the BRICS summit.
As chapter 7 explores in detail, GPT diffusion theory suggests that Xi, along with other leading policymakers and thinkers in both the United States and China, has learned the wrong lessons from previous industrial revolutions. If the IR-4 reshapes the economic power balance, the impact will materialize through a protracted period during which a GPT, such as AI, acquires a variety of uses in a wide range of productive processes. GPT skill infrastructure, not the flashy efforts to secure the high ground in innovation, will decide which nation owns the future in the IR-4.
Beyond power transitions, Technology and the Rise of Great Powers serves as a template for studying the politics of emerging technologies. An enduring dilemma is that scholars either assign too much weight to technological change or underestimate the effects of new technologies.52 Approaches that emphasize the social shaping of technology neglect that not all technologies are created equal, whereas technologically deterministic approaches discount the influence of political factors on technological development. By first distinguishing GPTs, together with their pattern of diffusion, from other technologies and technological trajectories, and then showing how social and political factors shape the pace and direction of GPT diffusion, my approach demonstrates a middle way forward.
Roadmap for the Book
The book proceeds as follows. Chapter 2 fleshes out the key differences between GPT diffusion theory and the LS-based account, as well as the case analysis procedures and selection strategy that allow me to systematically evaluate these two causal mechanisms. The bulk of the evidence follows in three case studies that trace how technological progress affected economic power transitions in the First, Second, and Third Industrial Revolutions.
The first two case studies, the IR-1 and IR-2, show that a gap in the adoption of GPTs, as opposed to monopoly profits from dominating LS innovations, was the crucial driver of an economic power transition. In both cases, the country that outpaced its industrial rivals made institutional adjustments to cultivate engineering skills related to the key GPT of the period. The IR-1 case, discussed in chapter 3, reveals that Britain was the most successful in fostering a wide pool of machinists who enabled the widespread diffusion of advances in iron machinery. In considering the IR-2 case, chapter 4 highlights how the United States surpassed Britain as the preeminent economic power by fostering a wide base of mechanical engineering talent to spread interchangeable manufacturing methods.
The IR-3 case, presented in chapter 5, demonstrates that technological revolutions do not necessarily always produce an economic power transition. The fact that Japan did not overtake the United States as the economic leader would provide disconfirming evidence of both the LS and GPT mechanisms, if the components of these mechanisms were present. In the case of the LS mechanism, Japan did dominate innovations in the IR-3’s leading sectors, including consumer electronics and semiconductor components. In contrast, the IR-3 does not discredit the GPT mechanism because Japan did not lead the United States in the diffusion of information and communications technology across a wide variety of economic sectors.
Chapter 6 uses large-n quantitative analysis to explore how GPT diffusion applies beyond great powers. Chapter 7 applies the GPT diffusion framework to the implications of modern technological breakthroughs for the US-China power balance. Focusing on AI technology as the next GPT that could transform the international balance of power, I explore the extent to which my findings generalize to the contemporary US-China case. I conclude in chapter 8 by underscoring the broader ramifications of the book.
2 GPT Diffusion Theory
HOW AND WHEN do technological changes affect the rise and fall of great powers? Specifically, how do significant technological breakthroughs result in differential rates of economic growth among great powers? International relations scholars have long observed that rounds of technological revolution lead to upheaval in global economic leadership, bringing about a power transition in the international system. However, few studies explore how this process occurs.
Those that do tend to fixate on the most dramatic aspects of technological change—the eureka moments and first implementations of radical inventions. Consequently, the standard account of technology-driven power transitions stresses a country’s ability to dominate innovation in leading sectors. By exploiting brief windows in which to monopolize profits in new industries, the country that dominates innovation in these sectors rises to become the world’s most productive economy. Explanations vary regarding why the benefits of leading sectors tend to accrue in certain nations. Some scholars argue that national systems of political economy that accommodate rising challengers can more readily accept and support new industries. Leading economies, by contrast, are victims of their past success, burdened by powerful vested interests that resist adaptation to disruptive technologies.1 Other studies point to more specific institutional factors that account for why some countries monopolize leading sectors, such as the degree of government centralization or industrial governance structures.2
An alternative explanation, based on the diffusion of general-purpose technologies (GPTs), draws attention to the less spectacular process by which fundamental innovations gradually diffuse throughout many industries. The rate and scope of diffusion is particularly relevant for GPTs, which are distinguished by their scope for continual improvement, broad applicability across many sectors, and synergies with other technological advances. Recognized by economists and historians as “engines of growth,” GPTs hold immense potential for boosting productivity.3 Realizing this promise, however, necessitates major structural changes across the technology systems linked to the GPT, including complementary innovations, organizational changes, and an upgrading of technical skills. Thus, GPTs lead to a productivity boost only after a “gradual and protracted process of diffusion into widespread use.”4 This is why more than five decades passed before key innovations in electricity, the quintessential GPT, significantly transformed manufacturing productivity.5
The process of GPT diffusion illuminates a pathway from technological change to power transition that diverges from the LS account (figure 2.1). Under the GPT mechanism, some great powers sustain economic growth at higher levels than their rivals do because, during a gradual process spanning decades, they more intensively adopt GPTs across a broad range of industries. This is analogous to a marathon run on a wide road. The LS mechanism, in contrast, specifies that one great power rises to economic leadership because it dominates innovations in a limited set of leading sectors and captures the accompanying monopoly profits. This is more like a sprint through a narrow running lane.
Why are some countries more successful at GPT diffusion? Building from scholarship arguing that a nation’s success in adapting to emerging technologies is determined by the fit between its institutions and the demands of evolving technologies, I argue that the GPT diffusion pathway informs the institutional adaptations crucial to success in technological revolutions.6 Unlike institutions oriented toward cornering profits in leading sectors, those optimized for GPT diffusion help standardize and spread novel best practices between the GPT sector and application sectors. Education and training systems that widen the base of engineering skills and knowledge linked to new GPTs, or what I call “GPT skill infrastructure,” are essential to all of these institutions.
FIGURE 2.1. Causal Diagrams for LS and GPT Mechanisms
The differences between these two theories of technological change and power transition are made clear when one country excels in the institutional competencies for LS product cycles but does not dominate GPT diffusion. Take, for example, chemical innovations and Germany’s economic rise in the late nineteenth century. Germany dominated major innovations in chemicals and captured nearly 90 percent of all global exports of synthetic dyestuffs.7 In line with the LS mechanism, this success was backed by Germany’s investments in building R&D labs and training doctoral students in chemistry, as well as a system of industrial organization that facilitated the rise of three chemical giants.8 Yet it was the United States that held an advantage in adopting basic chemical processes across many industries. As expected by GPT diffusion theory, the United States held institutional advantages in widening the base of engineering skills and knowledge necessary for chemicalization on a wide scale.9 This is when the ordinary tweakers and the implementers come to the fore, and the star scientists and inventors recede to the background.10
The rest of this chapter fleshes out my theoretical framework. It first clarifies the outcome I seek to explain: an economic power transition in which one great power becomes the economic leader by sustaining productivity growth at higher levels than its rivals. The starting point of my argument is that the diffusion of GPTs is central to the relationship between technological change and productivity leadership. This chapter explicates this argument by justifying the emphasis on both GPTs and diffusion, highlighting the differences between the GPT and LS mechanisms. It then extends the analysis to the institutional competencies that synergize with GPT trajectories. From the rich set of technology-institution interactions identified by evolutionary economists and comparative institutionalists, I justify my focus on institutions that enable countries to widen the skill base required to spread GPTs across industries. After differentiating my argument from alternative explanations, the chapter closes with a description of the book’s research methodology.11
The Outcome: Long-Term Economic Growth Differentials and Power Transitions
Power transitions are to the international system as earthquakes are to the geological landscape. Shifts in the relative power of leading nations send shock waves throughout the international system. What often follows is conflict, the most devastating form of which is a war waged by coalitions of great powers for hegemony over the globe.12 Beyond heightened risks of conflict, the aftershocks of power transitions reverberate in the architecture of the international order as victorious powers remake international institutions in their own images.13
While the power transition literature largely tackles the consequences of power transitions, I treat the rise and fall of great powers as the outcome to be explained. This follows David Baldwin’s instruction for international relations scholars “to devote more attention to treating power as a dependent variable and less to treating it as an independent variable.”14 Specifically, I explore the causes of “economic power transitions,” in which one great power sustains economic growth rates at higher levels than its rivals.15
It might not be obvious, at first glance, why I focus on economic power. After all, power is a multidimensional, contested concept that comes in many other forms. The salience of certain power resources depends on the context in which a country draws upon them to exert influence.16 For my purposes, differentials in economic growth are the most relevant considerations for intensifying hegemonic rivalry. An extensive literature has demonstrated that changes in relative economic growth often precede hegemonic wars.17
Moreover, changes in global political and military leadership often follow shifts in economic leadership. As the most fungible mode of power, economic strength undergirds a nation’s influence in global politics and its military capabilities.18 The outcomes of interstate conflicts bear out that economic and productive capacity is the foundation of military power.19 Paul Kennedy concludes that
all of the major shifts in the world’s military-power balances have followed alterations in the productive balances … the rising and falling of the various empires and states in the international system has been confirmed by the outcomes of the major Great Power wars, where victory has always gone to the side with the greatest material resources.20
How does one identify if or when an economic power transition occurs? Phrased differently, how many years does a great power need to lead its rivals in economic growth rates? How large does that gap have to reach? Throughout this book, I judge whether an economic power transition has occurred based on one great power attaining a lead in overall economic productivity over its rivals by sustaining higher levels of productivity growth rates.21 Productivity growth ensures that efficient and sustainable processes are fueling growth in total economic output. Additionally, productivity is the most important determinant of economic growth in the long run, which is the appropriate time horizon for understanding power transitions. “Productivity isn’t everything, but in the long run it is almost everything,” states Nobel Prize–winning economist Paul Krugman.22
Alternative conceptualizations of economic power cannot capture how effectively a country translates technological advance into national economic growth. Theories of geo-economics, for instance, highlight a state’s balance of trade in certain technologically advanced industries.23 Other studies emphasize a state’s share of world-leading firms.24 National rates of innovation, while more inclusive, measure the generation of novel technologies but not diffusion across commercial applications, thereby neglecting the ultimate impact of technological change.25 Compared to these indicators, which account for only a small portion of the value-added activities in the economy, productivity provides a more comprehensive measure of economic leadership.26
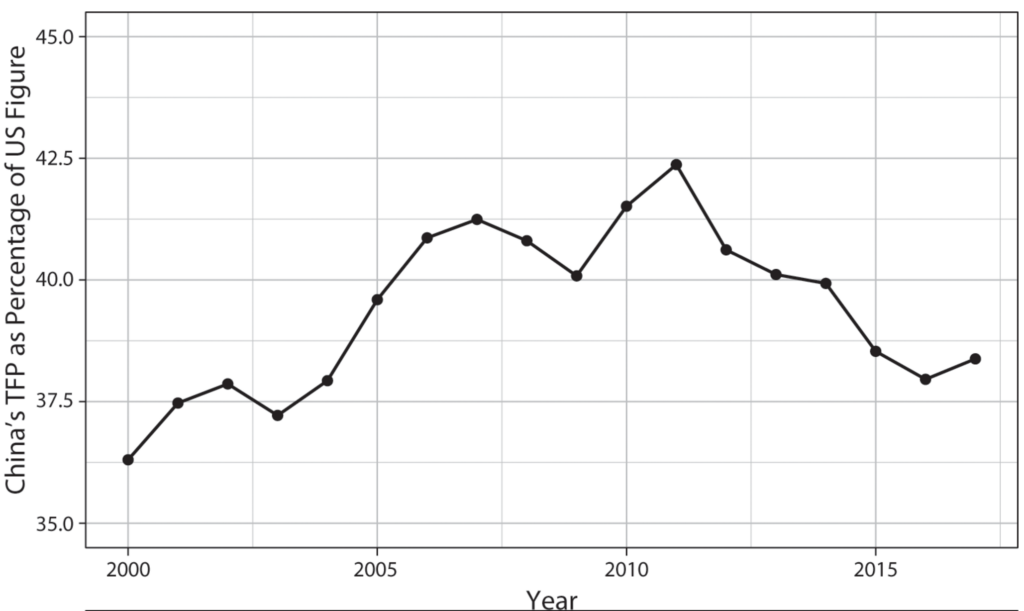
This focus on productivity is supported by recent work on power measurement, which questions measures of power resources based on economic size. Without accounting for economic efficiency, solely relying on measures of gross economic and industrial output provides a distorted view of the balance of power, particularly where one side is populous but poor.27 If national power was measured by GDP alone, China was the world’s most powerful country during the first industrial revolution. However, China’s economy was far from the productivity frontier. In fact, as the introduction chapter spotlighted, the view that China fell behind the West because it could not capitalize on productivity-boosting technological breakthroughs is firmly entrenched in the minds of leading Chinese policymakers and thinkers.
Lastly, it is important to clarify that I limit my analysis of productivity differentials to great powers.28 In some measures of productivity, other countries may rank highly or even outrank the countries I study in my cases. In the current period, Switzerland and other countries have higher GDP per capita than the United States; before World War I, Australia was the world leader in productivity, as measured by GDP per labor-hour.29 However, smaller powers like pre–World War I Australia and present-day Switzerland are excluded from my study of economic power transitions, as they lack the baseline economic and population size to be great powers.30
There is no exact line that distinguishes great powers from other countries.31 Kennedy’s seminal text The Rise and Fall of the Great Powers, for instance, has been challenged for not providing a precise definition of great power.32 Fortunately, across all the case studies in this book, there is substantial agreement on the great powers of the period. According to one measure of the distribution of power resources, which spans 1816 to 2012 and incorporates both economic size and efficiency, all the countries I study rank among the top six at the beginning of the case.33
The Diffusion of GPTs
Scholars often gravitate to technological change as the source of a power transition in which the mantle of industrial preeminence changes hands. However, there is less clarity over the process by which technical breakthroughs translate into this power shift among countries at the technological frontier. I argue that the diffusion of GPTs is the key to this mechanism. In this section, I first outline why my theory privileges GPTs over other types of technology. I then set forth why diffusion should be prioritized over other phases of technological change, especially innovation. Finally, I position GPT diffusion theory against the leading sector (LS) model, which is the standard explanation in the international relations literature.
Why GPTs?
Not all technologies are created equal. When assessed on their potential to transform the productivity of nations, some technical advances, such as the electric dynamo, rank higher than others, such as an improved sleeping bag. My theory gives pride of place to GPTs, such as electricity and the steam engine, which have historically generated waves of economy-wide productivity growth.34 Assessed on their own merits alone, even the most transformative technological changes do not tip the scale far enough to significantly affect aggregate economic productivity.35 GPTs are different because their impact on productivity comes from accumulated improvements across a wide range of complementary sectors; that is, they cannot be judged on their own merits alone.
Recognized by economists and economic historians as “engines of growth,” GPTs are defined by three characteristics.36 First, they offer great potential for continual improvement. While all technologies offer some scope for improvement, a GPT “has implicit in it a major research program for improvements, adaptations, and modifications.”37 Second, GPTs acquire pervasiveness. As a GPT evolves, it finds a “wide variety of uses” and a “wide range of uses.”38 The former refers to the diversity of a GPT’s use cases, while the latter alludes to the breadth of industries and individuals using a GPT.39 Third, GPTs have strong technological complementarities. In other words, the benefits from innovations in GPTs come from how other linked technologies are changed in response and cannot be modeled from a mere reduction in the costs of inputs to the existing production function. For example, the overall energy efficiency gains from merely replacing a steam engine with an electric motor were minimal; the major benefits from factory electrification came from electric “unit drive,” which enabled machines to be driven individually by electric motors, and a radical redesign of plants.40
Taken together, these characteristics suggest that the full impact of a GPT materializes via an “extended trajectory” that differs from those associated with other technologies. Economic historian Paul David explains:
We can recognize the emergence of an extended trajectory of incremental technical improvements, the gradual and protracted process of diffusion into widespread use, and the confluence with other streams of technological innovation, all of which are interdependent features of the dynamic process through which a general purpose engine acquires a broad domain of specific applications.41
For example, the first dynamo for industrial application was introduced in the 1870s, but the major boost of electricity to overall manufacturing productivity did not occur until the 1920s. Like other GPT trajectories, electrification required a protracted process of workforce skill adjustments, organizational adaptations, such as changes in factory layout, and complementary innovations like the steam turbine, which enabled central power generation in the form of utilities.42 To track the full impact of these engines of growth, one must travel the long roads of their diffusion.
Why Diffusion?
All technological trajectories can be divided into a phase when the technology is incubated and then first introduced as a viable commercial application (“innovation”) and a phase when the innovation spreads through a population of potential users, both nationally and internationally (“diffusion”).43 Recognizing this commonly accepted distinction, other studies of the scientific and technological capabilities of nations primarily focus on innovation.44 I depart from other works by giving priority to diffusion, since that is the phase of technological change most significant for GPTs.45
Undeniably, the activities and conditions that produce innovation can also spur diffusion.46 A firm’s ability to conduct breakthrough R&D does not just create new knowledge but also boosts its capacity to assimilate innovations from external sources (“absorptive capacity”).47 Faced with an ever-shifting technological frontier, building competency at producing new innovations gives a firm the requisite prior knowledge for identifying and commercializing external innovations. Other studies extend these insights beyond firms to regional and national systems.48 In order to absorb and diffuse technological advances first incubated elsewhere, they argue, nations must invest in a certain level of innovative activities.
This connection between innovation capacity and absorptive capacity could question the GPT mechanism’s attention to diffusion. Possibly, a country’s lead in GPT innovation could also translate directly into a relative advantage in GPT diffusion.49 Scholarship on the agglomeration benefits of innovation hot spots, such as Silicon Valley, support this case to some extent. Empirical analyses of patent citations indicate that knowledge spillovers from GPTs tend to cluster within a geographic region.50 In the case of electricity, Robert Fox and Anna Guagnini underscore that it was easier for countries with firms at the forefront of electrical innovation to embrace electric power at scale. The interconnections between the “learning by doing” gained on the job in these leading firms and academic labs separated nations in the “fast lane” and “slow lane” of electrification.51
Being the first to pioneer new technologies could benefit a state’s capacity to absorb and diffuse GPTs, but it is not determinative. A country’s absorptive capacity also depends on many other factors, including institutions for technology transfer, human capital endowments, openness to trade, and information and communication infrastructure.52 Sometimes the “advantages of backwardness” allow laggard states to adopt new technologies faster than the states that pioneer such advances.53 In theory and practice, a country’s ability to generate fundamental, new-to-the-world innovations can widely diverge from its ability to diffuse such advances.
This potential divergence is especially relevant for advanced economies, which encompass the great powers that are the subject of this research. Although innovation-centered explanations do well at sorting the advantages of technological breakthroughs to countries at the technological frontier compared to those trying to catch up, they are less effective at differentiating among advanced economies. As supported by a wealth of econometric research, divergences in the long-term economic growth of countries at the technology frontier are shaped more by imitation than innovation.54 These advanced countries have firms that can quickly copy or license innovations; first mover advantages from innovations are thus limited even in industries, like pharmaceuticals, that enforce intellectual property rights most strictly.55 Nevertheless, advanced countries that are evenly matched in their capacity for radical innovation can undertake vastly different growth trajectories in the wake of technological revolutions. Differences in diffusion pathways are central to explaining this puzzle.
This diffusion-centered approach is especially well suited for GPTs. Since GPTs entail gradual evolution into widespread use, there is a longer window for competitors to adopt GPTs more intensively than the leading innovation center. In other technologies, first-mover benefits from pioneering initial breakthroughs are more significant. For instance, leadership in the innovation of electric power technologies was fiercely contested among the industrial powers. The United States, Germany, Great Britain, and France all built their first central power stations within a span of three years (1882–1884), their first electric trams within a span of nine years (1887–1896), and their first three-phase AC power systems within a span of eight years (1891–1899).56 However, the United States clearly led in the diffusion of these systems: by 1912, its electricity production per capita had more than doubled that of Germany, its closest competitor.57 Thus, while most countries at the technological frontier will be able to compete in the production and innovation of GPTs, the hardest hurdles in the GPT trajectory are in the diffusion phase.
GPT Diffusion and LS Product Cycles
GPT diffusion challenges the LS-based account of how technological change drives power transitions. The standard explanation in the international relations literature emphasizes a country’s dominance in leading sectors, defined as new industries that experience rapid growth on the back of new technologies.58 Cotton textiles, steel, chemicals, and the automobile industry form a “classic sequence” of “great leading sectors,” developed initially by economist Walt Rostow and later adapted by political scientists.59 Under the LS mechanism, a country’s ability to maintain a monopoly on innovation in these emerging industries determines the rise and fall of lead economies.60
This model of technological change and power transition builds on the international product life cycle, a concept pioneered by Raymond Vernon. Constructed to explain patterns of international trade, the cycle begins with a product innovation and subsequent sales growth in the domestic market. Once the domestic market is saturated, the new product is exported to foreign markets. Over time, production shifts to these markets, as the original innovating country loses its comparative advantage.61
LS-based studies frequently invoke the product cycle model.62 Analyzing the effects of leading sectors on the structure of the international system, Gilpin states, “Every state, rightly, or wrongly, wants to be as close as possible to the innovative end of ‘the product cycle.’ ”63 One scholar described Gilpin’s US Power and the Multinational Corporation, one of the first texts that outlines the LS mechanism, as “[having] drawn on the concept of the product cycle, expanded it into the concept of the growth and decline of entire national economies, and analyzed the relations between this economic cycle, national power, and international politics.”64
The product cycle’s assumptions illuminate the differences between the GPT and LS mechanisms along three key dimensions. In the first stage of the product cycle, a firm generates the initial product innovation and profits from sales in the domestic market before saturation. Extending this model to national economies, the LS mechanism emphasizes the clustering of LS innovations and attendant monopoly profits in a single nation.65 “The extent of national success that we have in mind is of the fairly extreme sort,” write George Modelski and William Thompson. “One national economy literally dominates the leading sector during its phase of high growth and is the primary beneficiary of the immediate profits.”66 The GPT trajectory, in contrast, places more value on where technologies are diffused than where an innovation is first pioneered.67 I refer to this dimension as the “phase of relative advantage.”
In the next stage, the product innovation spreads to global markets and the technology gradually diffuses to foreign competitors. Monopoly rents associated with a product innovation dissipate as production becomes routinized and transfers to other countries.68 Mirroring this logic, Modelski and Thompson write, “[Leading sectors] bestow the benefits of monopoly profits on the pioneer until diffusion and imitation transform industries that were once considered radically innovative into fairly routine and widespread components of the world economy.”69 Thompson also states that “the sector’s impact on growth tends to be disproportionate in its early stages of development.”70
The GPT trajectory assumes a different impact timeframe. The more wide-ranging the potential applications of a technology, the longer the lag between its initial emergence and its ultimate economic impact. This explains why the envisioned transformative impact of GPTs does not appear straightaway in the productivity statistics.71 Time for complementary innovations, organizational restructuring, and institutional adjustments such as human capital formation is needed before the full impact of a GPT can be known. It is precisely the period when diffusion transforms radical innovations into routine components of the economy—the stage when the causal effects of leading sectors are expected to dissipate—that generates the productivity gap between nations.
The product cycle also reveals differences between the LS and GPT mechanisms regarding the “breadth of growth.” Like the product cycle’s focus on an innovation’s life cycle within a singular industry, the LS mechanism emphasizes the contributions of a limited number of new industries to economic growth in a particular period. GPT-fueled productivity growth, on the other hand, is dispersed across a broad range of industries.72 Table 2.1 specifies how LS product cycles differ from GPT diffusion along the three dimensions outlined here. As the following section will show, the differences in these two technological trajectories shape the institutional factors that are most important for national success in adapting to periods of technological revolution.
Table 2.1 Two Mechanisms of Technological Change and Power Transitions
Mechanisms
Impact Timeframe
Phase of Relative Advantage
Breadth of Growth
Institutional Complements
LS product cycles
Lopsided in early stages
Monopoly on innovation
Concentrated
Deepen skill base in LS innovations
GPT diffusion
Lopsided in later stages
Edge in diffusion
Dispersed
Widen skill base in spreading GPTs
While I have highlighted the differences between GPT diffusion and LS product cycles, it is important to recognize that there are similarities between the two pathways.73 Some scholars, for example, associate leading sectors with broad spillovers across economic sectors.74 In addition, lists of leading sectors and lists of GPTs sometimes overlap, as evidenced by the fact that electricity is a consensus inclusion on both lists. Moreover, both explanations begin with the same premise: to fully uncover the dynamics of technology-driven power transitions, it is essential to specify which new technologies are the key drivers of economic growth in a particular time window.75
At the same time, these resemblances should not be overstated. Many classic leading sectors do not have general-purpose applications. For instance, cotton textiles and automobiles both feature on Rostow’s series of leading sectors, and they are studied as leading sectors because each has “been the largest industry for several major industrial nations in the West at one time or another.”76 Although these were certainly both fast-growing large industries, the underlying technological advances do not fulfill the characteristics of GPTs. In addition, many of the GPTs I examine do not qualify as leading sectors. The machine tool industry in the mid-nineteenth century, for instance, was not a new industry, and it was never even close to being the largest industry in any of the major economies. Most importantly, though the GPT and LS mechanisms sometimes point to similar technological changes, they present very different understandings of how revolutionary technologies bring about an economic power transition. As the next section reveals, these differences also map onto varied institutional adaptations.
GPT Skill Infrastructure
New technologies agitate existing institutional patterns.77 They appeal for government support, generate new collective interests in the form of technical societies, and induce organizations to train people in relevant fields. If institutional environments are slow or fail to adapt, the development of emerging technologies is hindered. As Gilpin articulates, a nation’s technological fitness is rooted in the “extent of the congruence” between its institutions and the demands of evolving technologies.78 This approach is rooted in a rich tradition of work on the coevolution of technology and institutions.79
Understanding the demands of GPTs helps filter which institutional factors are most salient for how technological revolutions bring about economic power transitions. Which institutional factors dictate disparities in GPT adoption among great powers? Specifically, I emphasize the role of education and training systems that broaden the base of engineering skills linked to a particular GPT. This set of institutions, which I call “GPT skill infrastructure,” is most crucial for facilitating the widespread adoption of a GPT.
To be sure, GPT diffusion is dependent on institutional adjustments beyond GPT skill infrastructure. Intellectual property regimes, industrial relations, financial institutions, and other institutional factors could affect GPT diffusion. Probing inter-industry differences in technology adoption, some studies find that less concentrated industry structures are positively linked to GPT adoption.80 I limit my analysis to institutions of skill formation because their effects permeate other institutional arrangements.81 GPT skill infrastructure provides a useful indicator for other institutions that standardize and spread the novel best practices associated with GPTs.82
It should also be noted that the institutional approach is one of three main categories of explanation for cross-country differences in economic performance over the long term.83 Other studies document the importance of geography and culture to persistent cross-country income differences.84 I prioritize institutional explanations for two reasons. First, natural experiments from certain historical settings, in which institutional divergence occurs but geographical and cultural factors are held constant, suggest that institutional differences are particularly influential sources of long-term economic growth differentials.85 Second, since LS-based accounts of power transitions also prioritize institutional adaptations to technological change, my approach provides a more level test of GPT diffusion against the standard explanation.86
One final note about limits to my argument’s scope. I do not investigate the deeper origins of why some countries are more effective than others at developing GPT skill infrastructure. Possibly, the intensity of political competition and the inclusiveness of political institutions influence the development of skill formation institutions.87 Other fruitful lines of inquiry stress the importance of government capacity to make intertemporal bargains and adopt long time horizons in making technology investments.88 It is worth noting that a necessary first step to productively exploring these underlying causes is to establish which types of technological trajectories and institutional adaptations are at work. For instance, LS product cycles may be more closely linked to mercantilist or state capitalist approaches that favor narrow interest groups, whereas, political systems that incorporate a broader group of stakeholders may better accommodate GPT diffusion pathways.
Institutions Fit for GPT Diffusion
If GPTs drive economic power transitions, which institutions fit best with their demands? Institutional adaptations for GPT diffusion must solve two problems. First, since the economic benefits of GPTs materialize through improvements across a broad range of industries, capturing these benefits requires extensive coordination between the GPT sector and numerous application sectors. Given the sheer scope of potential applications, it is infeasible for firms in the GPT sector to commercialize the technology on their own, as the necessary complementary assets are embedded with different firms and industries.89 In the AI domain, as one example of a potential GPT, firms that develop general machine learning algorithms will not have access to all the industry-specific data needed to fine-tune those algorithms to particular application scenarios. Thus, coordination between the GPT sector and other organizations that provide complementary capital and skills, such as academia and competitor firms, is crucial. In contrast, for technologies that are not general-purpose, this type of coordination is less conducive and could even be detrimental to a nation’s competitive advantage, as the innovating firm could leak its technical secrets.90
Second, GPTs pose demanding conditions for human capital adjustments. In describing the connection between skill formation and technological fitness, scholars often delineate between general skills and industry-specific skills. According to this perspective, skill formation institutions that optimize for the former are more conducive to technological domains characterized by radical innovation, while institutions that optimize for the latter are more favorable for domains marked by incremental innovation.91 GPT diffusion entails both types of skill formation. The skills must be specific to a rapidly changing GPT domain but also broad enough to enable a GPT’s advance across many industries.92 Strong linkages between R&D-intensive organizations at the technological frontier and application areas far from the frontier also play a key role in GPT diffusion. This draws attention to the interactions between researchers who produce innovations and technicians who help absorb them into specific contexts.93
Education and training systems that foster relevant engineering skills for a GPT, or what I call GPT skill infrastructure, address both constraints. Engineering talent fulfills the need for skills that are rooted in a GPT yet sufficiently flexible to implement GPT advances in a wide range of sectors. Broadening the base of engineering knowledge also helps standardize best practices with GPTs, thereby coordinating information flows between the GPT sector and application sectors. Standardization fosters GPT diffusion by committing application sectors to specific technological trajectories and encouraging complementary innovations.94 This unlocks the horizontal spillovers associated with GPTs.95
Indeed, distinct engineering specializations have emerged in the wake of a new GPT. New disciplines, such as chemical engineering and electrical engineering, have proved essential in widening knowledge bases in the wake of a new GPT.96 Computer science, another engineering-oriented field, was central to US leadership in the information revolution.97 These professions developed alongside new technical societies—ranging from the American Society of Mechanical Engineers to the Internet Engineering Task Force—that formulated and disseminated guidelines and benchmarks for GPT development.98
Clearly, the features of GPT skill infrastructure have changed over time. Whereas informal associations systematized the skills crucial for mechanization in the eighteenth century, formal higher education has become increasingly integral to computerization in the twenty-first century.99 Some evidence suggests that computers and other technologies are skill-biased, in the sense that they favor workers with more years of schooling.100 These trends complicate but do not undercut the concept of GPT skill infrastructure. Regardless of the extent of formal training, all configurations of GPT skill infrastructure perform the same function: to widen the pool of engineering skills and knowledge associated with a GPT. This can take place in universities as well as in informal associations, provided these settings train engineers and facilitate the flow of engineering knowledge between knowledge-creation centers and application sectors.101
Which Institutions Matter?
The institutional competencies for exploiting LS product cycles are different. Historical analysis informed by this frame highlights heroic inventors like James Watt and pioneering research labs at large companies.102 Studying which countries benefited most from emerging technologies over the past two centuries, Herbert Kitschelt prioritizes the match between the properties of new technologies and sectoral governance structures. Under his framework, for example, tightly coupled technological systems with high causal complexity, such as nuclear power systems and aerospace platforms, are more likely to flourish in countries that allow for extensive state support.103 In other studies, the key institutional factors behind success in LS product cycles are education systems that subsidize scientific training and R&D facilities in new industries.104
These approaches equate technological leadership with a state’s success in capturing market shares and monopoly profits in new industries.105 In short, they use LS product cycles as the filter for which institutional variables matter. Existing scholarship lacks an institutional explanation for why some great powers are more successful at GPT diffusion.
Competing interpretations of technological leadership in chemicals during the late nineteenth century crystallize these differences. Based on the LS template, the standard account accredits Germany’s dominance in the chemical industry—as represented by its control over 90 percent of global production of synthetic dyes—to its investments in scientific research and highly skilled chemists.106 Germany’s dynamism in this leading sector is taken to explain its overall industrial dominance.107
GPT diffusion spotlights a different relationship between technological change and institutional adaptation. The focus turns toward institutions that complemented the extension of chemical processes to a wide range of industries beyond synthetic dye, such as food production, metals, and textiles. Under the GPT mechanism, the United States, not Germany, achieved leadership in chemicals because it first institutionalized chemical engineering as a discipline. Despite its disadvantages in synthetic dye production and chemical research, the United States was more effective in broadening the base of chemical engineering talent and coordinating information flows between fundamental breakthroughs and industrial applications.108
It is important to note that some parts of the GPT and LS mechanisms can coexist without conflict. A state’s capacity to pioneer new technologies can correlate with its capacity to absorb and diffuse GPTs. Countries that are home to cutting-edge R&D infrastructure may also be fertile ground for education systems that widen the pool of GPT-linked engineering skills. However, these aspects of the LS mechanism are not necessary for the GPT mechanism to operate. In accordance with GPT diffusion theory, a state can capitalize on GPTs to become the most powerful economy without monopolizing LS innovation.
Moreover, other dimensions of these two mechanisms directly conflict. When it comes to impact timeframe and breadth of growth, the GPT and LS mechanisms advance opposing expectations. Institutions suited for GPT diffusion can diverge from those optimized for creating new-to-the-world innovations. Research on human capital and long-term growth separates the effects of engineering capacity, which is commonly tied to adoptive activities, and other forms of human capital that are more often connected to inventive activities.109 This divergence can also be seen in debates over the effects of competition on technological activity. On the one hand, Joseph Schumpeter and others have argued that monopoly structures incentivize more R&D activity because the monopolists can appropriate all the gains from technological innovation.110 On the other hand, empirical work demonstrates that more competitive market structures increase the rate of technological adoption across firms.111 Thus, while there is some overlap between these two mechanisms, they can still be set against each other in a way that improves our understanding of technological revolutions and power transitions.
This theoretical framework differs from related work on the political economy of technological change.112 Scholars attribute the international competitiveness of nations to broader institutional contexts, including democracy, national innovation systems, and property rights enforcement.113 Since this book is limited to the study of shifts in productivity leadership at the technological frontier, many of these factors, such as those related to basic infrastructure and property rights, will not explain differences among technologically advanced nations.
In addition, most of the institutional theories put forth to explain the productivity of nations are technology-agnostic, in that they treat all forms of technological change equally. To borrow language from a former chairman of the US Council of Economic Advisers, they do not differentiate between an innovation in potato chips and an innovation in microchips.114 In contrast, I am specific about GPTs as the sources of shifts in competitiveness at the technological frontier.
Other theories identify key technologies but leave institutional factors at a high level of abstraction. Some scholars, for instance, posit that the lead economy’s monopoly on leading-sector innovation eventually erodes because of “ubiquitous institutional rigidities.”115 Unencumbered by the vested interests that resist disruptive technologies, rising challengers inevitably overtake established powers. Because these explanations are underspecified, they cannot account for cases where rich economies expand their lead or where poorer countries do not catch up.116
When interpreting great power competition at the technological frontier, adjudicating between the GPT and LS mechanisms represents a choice between two different visions. The latter prioritizes being the first country to introduce novel technologies, whereas the former places more value on disseminating and transforming innovations after their inception. In sum, industrial competition among great powers is not a sprint to determine which one can create the most brilliant Silicon Valley; it is a marathon won by the country that can cultivate the closest connections between its Silicon Valleys and its Iowa Citys.
Alternative Explanations
Although I primarily set GPT diffusion theory against the LS model, I also consider two other prominent explanations that make specific claims about how technological breakthroughs differentially advantage leading economies. Crucially, these two lines of thinking could account for differences in GPT diffusion, nullifying the import of GPT skill infrastructure.
Threat-Based Arguments
According to one school of thought, international security threats motivate states to invest in science and technology.117 When confronted with more threatening geopolitical landscapes, states are more incentivized to break down status quo interests and build institutions conducive to technological innovation.118 Militaries hold outsized influence in these accounts. For example, Vernon Ruttan argues that military investment, mobilized against war or the threat of war, fueled commercial advances in six technologies designated as GPTs.119 Studies of the success of the United States and Japan with emerging technologies also stress interconnections between military and civilian technological development.120 I group these related arguments under the category of threat-based theories.
Related explanations link technological progress with the balance of external threats and domestic roadblocks. Mark Taylor’s “creative insecurity” theory describes how external economic and military pressures permit governments to break from status quo interest groups and promote technological innovation. He argues that the difference between a nation’s external threats and its internal rivalries determines its propensity for innovation: the greater the difference, the greater the national innovation rate.121 Similarly, “systemic vulnerability” theory emphasizes the influence of external security and domestic pressures on the will of leaders to invest in institutions conducive to innovation, as well as the effect of “veto players” on their ability to do so.122
Certainly, external threats could impel states to invest more in GPTs, and military investment can help bring forth new GPTs; however, there are several issues with adapting threat-based theories to explain differences in GPT diffusion across great powers. First, threat-based arguments tend to focus on the initial incubation of GPTs, as opposed to the gradual spread of GPTs throughout a national economy. During the latter phase, a great deal of evidence suggests that civilian and military needs can greatly conflict.123 Besides, some GPTs, such as electricity in the United States, developed without substantial military investment. Since other civilian institutions could fill in as strong sources of demand for GPTs, military procurement may not be necessary for spurring GPT diffusion. Institutional adjustments to GPTs therefore can be motivated by factors other than threats. Ultimately, to further probe these points of clash, the impact of security threats and military investment must be traced within the historical cases.
Varieties of Capitalism
The “varieties of capitalism” (VoC) explanation highlights differences among developed democracies in labor markets, industrial organization, and interfirm relations and separates them into coordinated market economies (CMEs) and liberal market economies (LMEs). VoC scholars argue that CMEs are more suited for incremental innovations because their thick intercorporate networks and protected labor markets favor gradual adoption of new technological advances. LMEs, in contrast, are more adept at radical innovation because their fluid labor markets and corporate organization make it easier for firms to reorganize themselves around disruptive technologies. Most relevant to GPT diffusion theory, VoC scholars argue that LMEs incentivize workers to acquire general skills, which are more accommodative to radical innovation, whereas CMEs support industry-specific training, which is more favorable for incremental innovation.124
It is possible that differences between market-based capitalism and strategically coordinated capitalism account for GPT diffusion gaps between nations. Based on the expectations of the VoC approach, LMEs should be more likely to generate innovations with the potential to become GPTs, and workers in LMEs should possess more general skills that could spread GPTs across firms.125 Examining the pattern of innovation during the information revolution, scholars find that the United States, an LME, concentrated on industries experiencing radical innovation, such as semiconductors and telecommunications, while Germany, a CME, specialized in domains characterized by incremental innovation, such as mechanical engineering and transport.126
Despite bringing vital attention to the diversity of skill formation institutions, VoC theory’s dichotomy between general and industry-specific skills does not dovetail with the skills demanded by specific GPTs.127 Cutting across this sometimes arbitrary distinction, the engineering skills highlighted in GPT diffusion theory are specific to a fast-evolving GPT field and general enough to transfer ideas from the GPT sector across various sectors. Software engineering skills, for instance, are portable across multiple industries, but their reach is not as ubiquitous as critical thinking skills or mathematics knowledge. To address similar gaps in skill classifications, many political economists have appealed for “a more fine-grained analysis of cross-national differences in the particular mix of jobs and qualifications that characterize different political economies.”128 In line with this move, GPT skill infrastructure stands in for institutions that supply the particular mix of jobs and qualifications for enabling GPT diffusion. The empirical analysis provides an opportunity to examine whether this approach should be preferred to the VoC explanation for understanding technology-driven power transitions.
Research Methodology
My evaluation of the GPT and LS mechanisms primarily relies on historical case studies, which allow for detailed exploration of the causal processes that connect technological change to economic power transitions. Employing congruence-analysis techniques, I select cases and assess the historical evidence in a way that ensures a fair and rigorous test of the relative explanatory power of the two mechanisms.129 This sets up a “three-cornered fight” among GPT diffusion theory, the rival LS-based explanation, and the set of empirical information.130
The universe of cases most useful for assessing the GPT and LS mechanisms are technological revolutions (cause) that produced an economic power transition (outcome) in the industrial period. Following guidance on testing competing mechanisms that prioritize typical cases where the cause and outcome are clearly present, I investigate the First Industrial Revolution (IR-1) and the Second Industrial Revolution (IR-2).131 Both cases featured clusters of disruptive technological advances, highlighted by some studies as “technological revolutions” or “technology waves.”132 They also saw economic power transitions, when one great power sustained growth rates at substantially higher levels than its rivals.133 I also study Japan’s challenge to American economic leadership—which ultimately failed—in the Third Industrial Revolution (IR-3). This deviant case can disconfirm mechanisms and help explain why they break down.134
These cases are highly crucial and relevant for testing the GPT mechanism against the LS mechanism. All three cases favor the latter in terms of background conditions and existing theoretical explanations. Scholarship has attributed shifts in economic power during this period to the rise of new leading sectors.135 Thus, if the empirical results support the GPT mechanism, then my findings would suggest a need for major modifications to our understanding of how technological revolutions affect the rise and fall of great powers. The qualitative analysis appendix provides further details on case selection, including the universe of cases, the justification for these cases as “most likely cases” for the LS mechanism, and relevant scope conditions.136
This overall approach adapts the methodology of process-tracing, often conducted at the individual or micro level, to macro-level mechanisms that involve structural factors and evolutionary interactions.137 Existing scholarship on diffusion mechanisms, which the GPT mechanism builds from, emphasizes the influence of macro-level processes. In these accounts the diffusion trajectory depends not just on the overall distribution of individual-level receptivity but also on structural and institutional features, such as the degree of interconnectedness in a population.138 This approach aligns with a view of mechanistic thinking that allows for mechanisms to be set at different levels of abstraction.139 As Tulia Falleti and Julia Lynch point out, “Micro-level mechanisms are no more fundamental than macro-level ones.”140
To judge the explanatory strength of the LS and GPT mechanisms, I employ within-case congruence tests and process-tracing principles to evaluate the predictions of the two theoretical approaches against the empirical record.141 In each historical case, I first trace how leading sectors and GPTs developed in the major economies, paying particular attention to adoption timeframes, the technological phase of relative advantage, and the breadth of growth—three dimensions that differentiate GPT diffusion from LS product cycles.142
For example, my assessment of the two mechanisms along the impact time-frame dimension follows consistent procedures in each case.143 To evaluate when certain technologies were most influential, I establish when they initially emerged (based on dates of key breakthroughs), when their associated industries were growing fastest, and when they diffused across a wide range of application sectors. When data are available, I estimate a GPT’s initial arrival date by also factoring in the point at which it reached a 1 percent adoption rate in the median sector.144 Industry growth rates, diffusion curves, and output trends all help measure the timeline along which technological breakthroughs substantially influenced the overall economy. The growth trajectory of each candidate GPT and LS is then set against a detailed timeline of when a major shift in productivity leadership occurs.
I then turn to the institutional factors that could explain why some countries were more successful in adapting to a technological revolution, with a focus on the institutions best suited to the demands of GPTs and leading sectors.145 If the GPT mechanism is operative, the state that attains economic leadership should have an advantage in institutions that broaden the base of engineering human capital and spread best practices linked to GPTs. Additional evidence of the GPT diffusion theory’s explanatory power would be that other countries had advantages in institutions that complement LS product cycles, such as scientific research infrastructure and sectoral governance structures.
These evaluation procedures are effective because I have organized the competing mechanisms “so that they are composed of the same number of diametrically opposite parts with observable implications that rule each other out.”146 This allows for evidence in favor of one explanation to be doubly decisive in that it also undermines the competing theory.147 In sum, each case study is structured around investigating a set of four standardized questions that correspond to the three dimensions of the LS and GPT mechanisms as well as the institutional complements to technological trajectories (table 2.2).148
table 2.2 Testable Propositions of the LS and GPT Mechanisms
Dimensions
Key Questions
LS Propositions
GPT Propositions
Impact timeframe
When do revolutionary technologies make their greatest marginal impact on the economic balance of power?
New industries make their greatest impact on growth differentials in early stages.
GPTs do not make a significant impact on growth differentials until multiple decades after emergence.
Key phase of relative advantage
Do monopoly profits from innovation or benefits from more successful diffusion drive growth differentials?
A state’s monopoly on innovation in leading sectors propels it to economic leadership.
A state’s success in widespread adoption of GPTs propels it to economic leadership.
Breadth of growth
What is the breadth of technology-driven growth?
Technological advances concentrated in a few leading sectors drive growth.
Technological advances dispersed across a broad range of GPT-linked industries drive growth.
Institutional complements
Which types of institutions are most advantageous for national success in technological revolutions?
Key institutional adaptations help a state capture market shares and monopoly profits in new industries.
Key institutional adaptations widen the base of engineering skills and knowledge for GPT diffusion.
In each case study, I consider alternative theories of technology-driven power transitions. Countless studies have examined the rise and fall of great powers. My aim is not to sort through all possible causes of one nation’s rise or another’s decline. Rather, I am probing the causal processes behind an established connection between technological advances in each industrial revolution and an economic power transition. The VoC framework and threat-based theories outline alternative explanations for how significant technological advances translated into growth differentials among great powers. Across all the cases, I assess whether they provide a better explanation for the historical case evidence than the GPT and LS mechanisms.
I also address case-specific confounding factors. For example, some scholars argue that abundant inputs of wood and metals induced the United States to embrace more machine-intensive technology in the IR-2, reasoning that Britain’s slower adoption of interchangeable parts manufacturing was an efficient choice given its natural resource constraints.149 For each case, I determine whether these types of circumstantial factors could nullify the validity of the GPT and LS mechanisms.
In tracing these mechanisms, I benefit from a wealth of empirical evidence on past industrial revolutions, which have been the subject of many interdisciplinary inquiries. Since the cases I study are well-traversed terrain, my research is primarily based on secondary sources.150 I rely on histories of technology and general economic histories to trace how and when technological breakthroughs affected economic power balances. Notably, my analysis takes advantage of the application of formal statistical and econometric methods to assess the impact of significant technological advances, part of the “cliometric revolution” in economic history.151 Some of these works have challenged the dominant narrative of previous industrial revolutions. For instance, Nick von Tunzelmann found that the steam engine made minimal contributions to British productivity growth before 1830, raising the issue that earlier accounts of British industrialization “tended to conflate the economic significance of the steam engine with its early diffusion.”152
I supplement these historical perspectives with primary sources. These include statistical series on industrial production, census statistics, discussions of engineers in contemporary trade journals, and firsthand accounts from commissions and study teams of cross-national differences in technology systems. In the absence of standardized measures of engineering education, archival evidence helps fill in details about GPT skill infrastructure for each of the cases. In the IR-1 case, I benefit from materials from the National Archives (United Kingdom), the British Newspaper Archive, and the University of Nottingham Libraries, Manuscripts, and Special Collections. My IR-2 case analysis relies on collections based at the Bodleian Library (United Kingdom), the Library of Congress (United States), and the University of Leipzig and on British diplomatic and consular reports.153 In the IR-3 case analysis, the Edward A. Feigenbaum Papers collection, held at Stanford University, helps inform US-Japan comparisons in computer science education.
My research also benefits greatly from new data on historical technological development. I take advantage of improved datasets, such as the Maddison Project Database.154 New ones, such as the Cross-Country Historical Adoption of Technology dataset, were also beneficial.155 Sometimes hype about exciting new technologies influences the perceptions of commentators and historians about the pace and extent of technology adoption. More granular data can help substantiate or cut through these narratives. Like the reassessments of the impact of previous technologies, these data were released after the publication of the field-defining works on technology and power transitions in international relations. Making extensive use of these sources therefore provides leverage to revise conventional understandings.
When assessing these two mechanisms, one of the main challenges is to identify the key technological changes to trace. I take a broad view of technology that encompasses not just technical designs but also organizational and managerial innovations.156 Concretely, I follow Harvey Brooks, a pioneer of the science and technology policy field, in defining technology as “knowledge of how to fulfill certain human purposes in a specifiable and reproducible way.”157 The LS and GPT mechanisms both call attention to the outsized import of particular technical breakthroughs and their interactions with social systems, but they differ on which ones are more important. Therefore, a deep and wide understanding of advances in hardware and organizational practices in each historical period is required to properly sort them by their potential to spark LS or GPT trajectories.
This task is complicated by substantial disagreements over which technologies are leading sectors and GPTs. Proposed lists of GPTs often conflict, raising questions about the criteria used for GPT selection.158 Reacting to the length of such lists, other scholars fear that “the [GPT] concept may be getting out of hand.”159 According to one review of eleven studies that identified past GPTs, twenty-six different innovations appeared on at least one list but only three appeared on all eleven.160
The LS concept is even more susceptible to these criticisms because the characteristics that define leading sectors are inconsistent across existing studies. Though most scholars agree that leading sectors are new industries that grow faster than the rest of the economy, there is marked disagreement on other criteria. Some scholars select leading sectors based on the criterion that they have been the largest industry in several major industrial nations for a period of time.161 Others emphasize that leading sectors attract significant investments in R&D.162 To illustrate this variability, I reviewed five key texts that analyze the effect of leading sectors on economic power transitions. Limiting the lists of proposed leading sectors to those that emerged during the three case study periods, I find that fifteen leading sectors appeared on at least one list and only two appeared on all five.163
My process for selecting leading sectors and GPTs to trace helps alleviate concerns that I cherry-pick the technologies that best fit my preferred explanation. In each historical case, most studies that explicitly identify leading sectors or GPTs agree on a few obvious GPTs and leading sectors. To ensure that I do not omit any GPTs, I consider all technologies singled out by at least two of five key texts that identify GPTs across multiple historical periods.164 I apply the same approach to LS selection, using the aforementioned list I compiled.
Following classification schemes that differentiate GPTs from “near-GPTs” and “multipurpose technologies,” I resolve many of the conflicts over what counts as a GPT or leading sector by referring to a set of defining criteria.165 For instance, while some accounts include the railroad and the automobile as GPTs, I do not analyze them as candidate GPTs because they lack a variety of uses.166 I support my choices with empirical methods for LS and GPT identification. To confirm certain leading sectors, I examine the rate of growth across various industry sectors. I also leverage recent studies that identify GPTs with patent-based indicators.167 Taken together, these procedures limit the risk of omitting certain technologies while guarding against GPT and LS concept creep.168 The qualitative analysis appendix outlines how I address other issues related to LS and GPT identification, including concerns about omitting important single-purpose technologies and scenarios when certain technological breakthroughs are linked to both LS and GPT trajectories.
These considerations underscore that taking stock of the key technological drivers is only the first step in the case analysis. To judge whether these breakthroughs actually brought about the impacts that are often claimed for them, it is important to carefully trace how these technologies evolved in close relation with societal systems.
As a complement to the historical case studies, this book’s research design includes a large-n quantitative analysis of the relationship between the breadth of software engineering skill formation institutions and computerization rates. This tests a key observable implication of GPT diffusion theory, using time-series cross-sectional data on nineteen advanced and emerging economies across three decades. I leave the more detailed description of the statistical methodology to chapter 6.
Summary
The technological fitness of nations is determined by how they adapt to the demands of new technical advances. I have developed a theory to explain how revolutionary technological breakthroughs affect the rise and fall of great powers. My approach is akin to that of an investigator tasked with figuring out why one ship sailed across the ocean faster than all the others. As though differentiating the winning ship’s route from possible sea-lanes in terms of trade wind conditions and course changes, I first contrast the GPT and LS trajectories with regard to the timing, phase, and breadth of technological change. Once the superior route has been mapped, attention turns to the attributes of the winning ship, such as its navigation equipment and sailors’ skills, that enabled it to take advantage of this fast lane across the ocean. In similar fashion, having set out the GPT trajectory as the superior route from technological revolution to economic leadership, my argument then highlights GPT skill infrastructure as the key institutional attribute that dictates which great power capitalizes best on this route.
3 The First Industrial Revolution and Britain’s Rise
FEW HISTORICAL EVENTS have shaken the world like the First Industrial Revolution (IR-1, 1780–1840). Extraordinary upheaval marked the contours and consequences of the IR-1. For the first time in history, productivity growth accelerated dramatically, allowing large numbers of people to experience sustained improvements in their living standards. Small towns transformed into large cities, new ideologies gathered momentum, and emerging economic and social classes reshaped the fabric of society. These changes reverberated in the international sphere, where the ramifications of the IR-1 included the transition to industrialized mass warfare, the decline of the absolutist state, and the birth of the modern international system.
Among these transformations, two phenomena stand out. The first is the remarkable technological progress that inaugurated the IR-1 period. Everything was changing in part because so many things were changing—water frames, steam engines, and puddling processes not least among them. The second is Britain’s rise to unrivaled economic leadership, during which it sustained productivity growth at higher levels than its rivals, France and the Netherlands. The following sections adjudicate the debates over the exact timeline of Britain’s industrialization, but there is no doubt that Britain, propelled by the IR-1, became the world’s most advanced economic power by the mid-nineteenth century.
No study of technological change and power transitions is complete without an account of the IR-1. For both the LS and GPT mechanisms, the IR-1 functions as a typical case that is held up as paradigmatic of technology-driven power transitions. The standard account in international relations scholarship attributes Britain’s industrial ascent to its dominance of innovation in the IR-1’s leading sectors, including cotton textiles, iron metallurgy, and steam power.1 Present-day scholarship and policy discussions often draw upon stylized views of the IR-1, analogizing present developments in information technology and biotechnology to the effects of steam power and cotton textiles in the industrial revolution.2
A deeper inquiry into the IR-1 and Britain’s economic rise challenges many of these conventional views. First, it reveals that general-purpose transformations linked to advances in iron metallurgy diffused widely enough to significantly affect economy-wide productivity only after 1815—a timeline that aligns with the period when Britain significantly outpaced its rivals in industrialization. Other prominent advances, including the steam engine, made only limited contributions to Britain’s rise to industrial prominence in this period owing to a prolonged period of gestation before widespread adoption. Second, the IR-1 case also demonstrates that it was Britain’s advantage in extending mechanization throughout the economy, not monopoly profits from innovations in cotton textiles, that proved crucial to its industrial ascendancy. Third, the historical data illustrate that the dispersion of mechanical innovations across many sectors fueled British productivity growth. Across these three dimensions, the IR-1 case matches the GPT trajectory better than the LS trajectory.
Since no country monopolized innovations in metalworking processes and Britain’s competitors could also absorb innovations from abroad, why did Britain gain the most from this GPT trajectory? In all countries, as technical advances surged forward, institutional adjustments raced to cultivate the skills required to keep pace. Importantly, France and the Netherlands were competitive with Britain—and even surpassed it in some respects—in scientific research infrastructure and education systems for training expert engineers. These institutional settings in France and the Netherlands, however, produced knowledge and skills that were rather divorced from practical applications.
Britain’s comparative advantage rested on another type of skill infrastructure. It depended less on heroic innovators like James Watt, the famed creator of the modern steam engine, and more on competent engineers who could build and maintain new technological systems, as well as make incremental adaptations to implement these systems in many different settings.3 As expected by GPT diffusion theory, Britain benefited from education systems that expanded the base of mechanically skilled engineers and disseminated knowledge of applied mechanics. Britain’s competitors could not match its system for cultivating a common technical language in applied mechanics that encouraged the knowledge exchanges between engineers and entrepreneurs needed for advancing mechanization from one industry to the next.
To trace these mechanisms, I gathered and sorted through a wealth of evidence on the IR-1. Historical accounts served as the foundational materials, including general economic histories of the IR-1, histories of influential technologies and industries like the steam engine and the iron industry, country-specific histories, and comparative histories of Britain, France, and the Netherlands. I also benefited from contemporary assessments of the IR-1’s institutional features provided by trade journals, proceedings of mechanics’ institutes, recruitment advertisements published in local newspapers, and essays by leading engineers. This evidence stems from archival materials at the British Newspaper Archive, the National Archives (United Kingdom), and the University of Nottingham Libraries, Manuscripts, and Special Collections. Triangulating a variety of sources, I endeavored to back up my claims with statistical evidence in the form of industrial output estimates, patenting rates, and detailed biographical information on British engineers.
The assessment of the GPT and LS mechanisms against historical evidence from the IR-1 proceeds as follows. To begin, the chapter reviews Britain’s rise to industrial preeminence, which is the outcome of the case. Next, it sorts the key technological breakthroughs of the period by their potential to drive two types of trajectories—LS product cycles and GPT diffusion. I then assess whether Britain’s rise in this period is better explained by the GPT or LS mechanism, tracing the development of candidate leading sectors and GPTs in terms of impact timeframe, phase of relative advantage, and breadth of growth. If the GPT trajectory holds for this period, there should be evidence that Britain was better equipped than its competitors in GPT skill infrastructure. Another section evaluates whether the historical data support this expectation. Before concluding the chapter, I address alternative factors and explanations.
A Power Transition: Britain’s Rise
When did Britain ascend to industrial hegemony? The broad outlines of the story are well known. Between the mid-eighteenth century and the mid-nineteenth century, the industrial revolution propelled Great Britain to global preeminence. Although Britain did not boast the world’s largest economy—China held that title during this period—it did capitalize on the technologies of the industrial revolution to become “the world’s most advanced productive power.”4 France and the Netherlands, its economic rivals, did not keep pace with Britain’s productivity growth.
While both the LS and GPT models agree that Britain established itself as the preeminent industrial power in this period, a clearer sense of when this shift occurred is essential for testing the explanatory power of the LS and GPT mechanisms during this period. One common view of Britain’s industrialization, brought to prominence by Rostow, depicts an accelerated takeoff into sustained growth. Rostow’s timeline dates this takeoff to the last two decades of the eighteenth century.5 In alignment with this periodization, some scholars writing in the LS tradition claim that Britain achieved its industrial ascent by the late eighteenth century.6
A different perspective, better supported by the evidence that follows, favors a delayed timeline for Britain’s ascent to industrial preeminence. Under this view, Britain did not sustain economic and productivity advances at levels substantially higher than its rivals until the 1820s and after. To clarify the chronology of Britain’s industrial ascent, the following sections survey three proxies for productivity leadership: GDP per capita, industrialization, and total factor productivity.
GDP PER-CAPITA INDICATORS
Trend lines in GDP per capita, a standard proxy for productivity, confirm the broad outlines of Britain’s rise. Evidence from the Maddison Project Database (MPD) points to the decades after 1800, not before, as the key transition period (figure 3.1).7 These trends come from the 2020 version of the MPD, which updates Angus Maddison’s data and incorporates new annual estimates of GDP per capita in the IR-1 period for France, the Netherlands, and the United Kingdom.8 In 1760, the Netherlands boasted the world’s highest per-capita income, approximately 35 percent higher than Britain’s.9 The Dutch held this lead for the rest of the eighteenth century through to 1808, when Britain first overtook the Netherlands in GDP per capita. By 1840, Britain’s GDP per capita was about 75 percent higher than that of France and about 10 percent ahead of that of the Netherlands.10
figure 3.1 Economic Power Transition in the IR-1. Source: Maddison Project Database, version 2020 (Bolt and van Zanden 2020).
It should be noted that GDP per-capita information for the early years of the IR-1 is sometimes missing or only partially available. For years prior to 1807, the MPD bases Dutch GDP per-capita estimates on data for just the Holland region, so the Dutch economy’s decline during this time could be an artifact of changes in data sources.11 At the same time, to ensure that the MPD data can be used to provide accurate information on historical patterns of economic growth and decline, researchers have made adjustments to partial data series and consulted experts to assess their representativeness.12 Furthermore, the Holland-based data in the early 1800s already indicated a decline in Dutch GDP per capita. Although data scarcity makes it difficult to mark out exactly when Britain’s GDP per capita surpassed that of the Netherlands, the MPD remains the best source for cross-country comparisons of national income in this period.
INDUSTRIALIZATION INDICATORS
Industrialization indicators depict a mixed picture of when Britain sustained leadership in economic efficiency. By one influential set of metrics compiled by economic historian Paul Bairoch, Britain’s per-capita industrialization levels had grown to 50 percent higher than those of France in 1800, from a position of near-equality with France and Belgium in 1750. For scholars who map the trajectories of great powers, these estimates have assumed a prominent role in shaping the timeline of British industrial ascendance.13 For instance, Paul Kennedy employs Bairoch’s estimates to argue that the industrial revolution transformed Britain into a different kind of world power.14
Further examination of Bairoch’s estimates qualifies their support for an accelerated timeline of Britain’s industrial ascendance. First, by limiting his definition of “industrial output” to manufacturing industry products, Bairoch excludes the contribution of notable sectors such as construction and mining, a distinction even he admits is “rather arbitrary.”15 Second, the gap between Britain and France in per-capita industrialization levels in 1800 still falls within the margin of error for Bairoch’s estimates.16
Moreover, a delayed timeframe is supported by industrialization measures that encompass more than the manufacturing industries. In 1700, the Netherlands had a substantially higher proportion of its population employed in industry (33 percent) compared to the United Kingdom (22 percent). In 1820, the proportion of people employed in UK industry had risen to 33 percent—higher than the Dutch corresponding rate of 28 percent.17 One expert on the pre-industrial revolution in Europe notes that the Netherlands was at least as industrialized as England, if not more so, throughout the eighteenth century.18 Lastly, aggregate industrial production trends map out a post-1815 surge in British industrialization, providing further evidence that Britain did not solidify its productivity advantage until decades into the nineteenth century.19
PRODUCTIVITY INDICATORS
Total factor productivity (TFP) indicators, which capture the efficiency by which production factors are converted into useful outputs, further back the delayed ascent story. As was the case with trends in aggregate industrial output, TFP growth in Britain did not take off until after 1820.20 In truth, British TFP growth was very modest throughout the eighteenth century, averaging less than 1 percent per year.21
While the paucity of reliable data on total factor productivity in France hinders cross-national comparisons in this period, some evidence suggests that Britain did not surpass the Netherlands in TFP until after 1800.22 Historian Robert Allen estimated TFP in agriculture by calculating the ratio between actual output per worker and the output per worker predicted by a country’s available agricultural population and land. On this metric for the year 1800, the Netherlands ranked higher than Britain and all other European nations.23 The Dutch also attained the highest absolute TFP in Europe for almost all of the eighteenth century.24
Which periodization of Britain’s industrial ascent better reflects the empirical evidence? On balance, measures of per-capita GDP, industrialization levels, and total factor productivity support a deferred timeline for Britain’s industrial rise. This clarification of when an economic power transition occurred during the IR-1 provides a stable target to test the competing LS and GPT mechanisms.
Key Technological Changes in the IR-1
Before evaluating the LS and GPT mechanisms in greater depth, the technological elements of the IR-1 must be further specified. Hargreaves’s spinning jenny (1764), Arkwright’s water frame (1769), Watt’s steam engine (1769), Cort’s puddling process (1784), and many other significant technical advances emerged during the First Industrial Revolution. The most likely sources of GPT and LS trajectories can be identified with guidance from existing work that calls attention to key technologies and accepted criteria for these two categories. Narrowing the assessment of these two mechanisms to a limited set of technologies makes for a more viable exercise.
Candidate Leading Sectors: Cotton Textiles, Iron, and Steam Power
A strong degree of consensus on the leading sectors that powered Britain’s rise in the IR-1 makes it relatively easy to identify three candidate sectors: cotton textiles, iron, and steam power.25 Among these, historians widely recognize the cotton textile industry as the original leading sector of the First Industrial Revolution.26 New inventions propelled the industry’s rapid growth, as its share of total value added to British industry rose from 2.6 percent in 1770 to 17 percent in 1801.27 In characterizing the significance of the cotton industry, Schumpeter went as far as to claim, “English industrial history can, in the epoch 1787–1842 … be almost resolved into the history of a single industry.”28
If the cotton textile industry places first in the canon of the IR-1’s leading sectors, then the iron industry follows close behind. In their account of Britain’s rise, Modelski and Thompson single out these two major industries, employing pig iron production and cotton consumption, as indicators for Britain’s leading sector growth rates.29 According to the traditional view of the IR-1, the iron and cotton industries were the only two that experienced “highly successful, rapidly diffused technical change” before the 1820s.30
I also evaluate the steam power industry as a third possible leading sector. A wide range of LS-based scholarship identifies steam power as one of the technological foundations of Britain’s leadership in the nineteenth century.31 Most of this literature labels only the steam engine itself as the leading sector, but since leading sectors are new industries, the steam engine–producing industry is the more precise understanding of the leading sector related to major advances in steam engine technology. Compared to the iron and cotton textile industries, it is much more uncertain whether the steam engine–producing industry, which experienced relatively slow growth in output and productivity, meets the analytical criteria for a leading sector during the IR-1 case.32 Still, I include the steam engine–producing industry as a potential leading sector, leaving it to the case analysis to further study its growth trajectory.
Candidate GPTs: Iron, Steam Engine, and the Factory System
Since the possible sources of GPT trajectories in the IR-1 are less established, I drew on previous studies that mapped out technological paradigms in this period to select three candidate GPTs: the steam engine, mechanization based on advances in iron machinery, and the factory system.33 As a possible source of GPT-style effects, the steam engine is a clear choice. Alongside electricity and (ICT) technology, it has been described as one of the “Big Three” GPTs, appearing in nearly all catalogs of GPTs.34 Here the emphasis is on the capacity of steam engines to transform a wide variety of industrial processes across many sectors, as opposed to the potential growth of the steam engine–producing industry.
Of the two paradigmatic industries of the IR-1, cotton and iron, the latter was a more plausible driver of GPT-style effects for Britain. As the demand for iron-made machinery grew, iron foundries development of new generations of machine tools, such as cylinder-boring machines, contributed to the creation of a mechanical engineering industry.35 This spurred the mechanization of production processes in a wide range of industries, including agriculture, food processing, printing, and textiles.36 Although both cotton textiles and iron were fast-growing industries, developments in iron better resembled a “motive branch” driving pervasive effects across the economy.37
In addition, the late eighteenth century saw the emergence of centralized factories, which significantly increased the scale of goods production. The factory system offered the potential to change the techniques of production across many industries. One widely cited classification scheme for GPTs picks out the factory system as an “organizational GPT” in the late eighteenth to early nineteenth century period.38 Other scholars describe this organizational innovation as “one of the most fundamental changes of ‘metabolism’ in the Industrial Revolution.”39
I also considered but ultimately decided against including developments in railroads as a candidate GPT. Among five core texts that classify GPTs across many historical periods, at least two highlighted the significance of the railroad to the IR-1.40 In my view, the railroad represented a disruptive advance, but it did not acquire the variety of uses to qualify as a GPT. Railways carried many types of freight and made new business models possible, but their function was limited to transport.41
Sources of LS and GPT Trajectories
Table 3.1 recaps the potential technological sources for both the GPT and LS mechanisms. It is important to clarify three points about the sorting process. First, it is notable but not surprising that the candidate leading sectors and GPTs draw from similar technological wellsprings. Both mechanisms agree that some inventions, like Cort’s puddling process for making wrought iron, mattered much more than others in terms of their impact on the economic balance of power.
Table 3.1 Key Sources of Technological Trajectories in the IR-1
Candidate Leading Sectors
Candidate GPTs
Cotton textile industry
Factory system
Iron industry
Mechanization
Steam engine–producing industry
Steam engine
Where the mechanisms separate is in how this process transpired. Cort’s puddling process and other ironmaking innovations, under the GPT model, are expected to follow an impact pathway characterized by three features: an extended lag before they affect productivity growth, the spread of mechanization across the economy, and widespread complementary innovations in many machine-using industries. Under the LS model, the same technological sources are expected to affect economic growth in a way that is lopsided in the early stages of development, fueled by monopolizing iron exports, and limited to technological innovations in the iron industry.
Second, it is still useful to classify three distinct candidate GPTs in the IR-1, despite the fact that developments in factory systems, mechanization, and steam engines were mutually reinforcing in many respects. Steam engines depended on the transition from hand-tool processes to machinery-based production systems; at the same time, the impact of steam engines on coal mining was to boost the iron industry, spurring mechanization. Yet a number of historians distinguish the expansion of mechanization in the British industrial revolution from transformations linked to the steam engine, arguing that the latter’s economic impact materialized much later than the former.42 Thus, while these candidate GPT trajectories are interconnected, it is still possible to locate various GPTs at different stages of their life cycle.
Third, not all of these technological changes necessarily had a decisive impact on Britain’s capacity to sustain higher productivity levels than its rivals during the period of interest. They are labeled as candidates for a reason. As this chapter will show, the steam engine did not achieve widespread diffusion until after Britain had already established economic leadership. When subjected to more rigorous empirical analysis, developments in some technologies may not track well with the proposed LS and GPT trajectories for this period.
GPT vs. LS Trajectories in the IR-1
Spelling out possible sources of technological trajectories in the IR-1 provides a bounded terrain for testing the validity of the GPT and LS mechanisms. By leveraging differences between the two mechanisms with respect to impact timeframe, phase of relative advantage, and breadth of growth, I derive three sets of opposing predictions for how technological changes translated into an economic power transition in this period. I then assess whether, and to what extent, the developments in the IR-1 supported these predictions.
OBSERVABLE IMPLICATIONS RELATED TO IMPACT TIMEFRAME
When did the revolutionary technologies of the IR-1 disrupt the economic balance of power? If the impact timeframe of the LS mechanism holds, then radical technical advances in the cotton textile, iron, and/or steam engine–producing industries should have substantially stimulated British economic growth shortly after the emergence of major technological advances in the 1760s and 1770s.43 Accordingly, scholars theorize that leading sectors propelled Great Britain to industrial superiority in the late eighteenth century.44 In line with this conception of a rapid timeline, Modelski and Thompson expect that the growth of two lead industries, cotton and iron, peaked in the 1780s.45
On the other hand, if the GPT mechanism was operational, the impact of major technological breakthroughs on Britain’s industrial ascent should have arrived on a more gradual timeline. Key advances tied to mechanization, steam power, and the factory system emerged in the 1770s and 1780s. Given that GPTs require a long period of delay before they diffuse and achieve widespread adoption, the candidate GPTs of the IR-1 should not have had substantial economy-wide repercussions until the early decades of the nineteenth century and after. I use the year 1815 as a rough cut-point to separate the accelerated impact timeframe of leading sectors from that of GPTs in this period.
OBSERVABLE IMPLICATIONS RELATED TO THE PHASE OF RELATIVE ADVANTAGE
The LS mechanism places high value on the innovation phase of technological change. Where major breakthroughs arise is key. Accordingly, Britain’s capacity to pioneer major technological advances should explain economic growth differentials in the IR-1. Concretely, the LS mechanism expects that Britain’s rise was fueled by its dominance of innovation in the cotton textile, iron, and steam engine–producing industries, as well as the resultant monopoly rents from exports in these sectors.
The GPT mechanism emphasizes a less-celebrated phase of technological change. Where innovations diffuse is key. Differentials in the rate and intensiveness of GPT adoption generate the gap between an ascending industrial leader and other competitors. The GPT mechanism suggests that Britain’s rise to industrial preeminence can be traced to its superior ability to diffuse generic technological changes across the economy.
OBSERVABLE IMPLICATIONS RELATED TO BREADTH OF GROWTH
The last set of observable implications relate to the breadth of growth during the IR-1. As illustrated in the descriptions here of the candidate leading sectors, many accounts attribute Britain’s industrial ascent to a narrow set of critical advances.46 In one of the first texts dedicated to the investigation of technology and international relations, William Ogburn declared, “The coming of the steam engine … is the variable which explains the increase of Britain as a power in the nineteenth century.”47 According to GPT diffusion theory, Britain’s rise to industrial preeminence came from the advance of GPTs through many linked sectors.
Taken together, these three sets of diverging predictions guide my assessment of the GPT mechanism against the LS mechanism. I make expectations specific to the IR-1 case by using the relevant information on particular technologies and the timeline of British industrialization. Table 3.2 lays out the specific, testable predictions that provide the framework of evaluation in the following sections.
Table 3.2 Testable Predictions for the IR-1 Case Analysis
Prediction 1: LS (impact timeframe)
The cotton textile, iron, and/or* steam engine–producing industries made a significant impact on Britain’s rise to industrial preeminence before 1815.
Prediction 1: GPT
Mechanization, the steam engine, and / or the factory system made a significant impact on Britain’s rise to industrial preeminence only after 1815.
Prediction 2: LS (relative advantage)
Innovations in cotton textile, iron, and/or the steam engine–producing industries were concentrated in Britain.British advantages in the production and exports of textiles, iron, and/or steam engines were crucial to its industrial superiority.
Prediction 2: GPT
Innovations in iron, the steam engine, and/or the factory system were not concentrated in Britain.
British advantages in the diffusion of mechanization, steam engines, and/or the factory system were crucial to its industrial superiority.
Prediction 3: LS (breadth of growth)
Productivity growth in Britain was limited to the cotton textile, iron, and/or steam engine – producing industries.
Prediction 3: GPT
Productivity growth in Britain was spread across a broad range of industries linked to mechanization, the steam engine, and/or the factory system.
* The operator “and/or” links all candidate leading sectors and GPTs because it could be the case that only some of these technologies drove the trajectories of the period.
Impact Timeframe: Delayed Surge vs. Fast Rise of British Industrialization
The painstaking reconstruction of temporal chronology is at the heart of tracing mechanisms. Tremendous technological changes occurred during this period, but when exactly did they make their mark on Britain’s industrial superiority? The period when significant technological innovations emerge often does not match up with the time when their impacts are felt. Unfortunately, when drawing lessons from the IR-1 on the effect of technology on international politics, scholars have conflated the overall significance of certain technologies with near-immediate impact.48 Establishing a clear timeline of when technological changes catalyzed a shift in productivity leadership during the IR-1 is therefore an important first step in comparing the LS and GPT mechanisms.
DIVERGING TIMELINES: COTTON VS. IRON
Time-series data on the output growth of twenty-six industries that accounted for around 60 percent of Britain’s industrial production help differentiate the growth schedules of the cotton textiles and iron industries. According to these data, the major upswing in British industrialization took place after 1815, when the aggregate growth trend increased from 2 percent to a peak of 3.8 percent by 1825. In line with the expectations of the LS model, the cotton textile industry grew exceptionally fast following major technological innovations in the 1760s, but from the 1780s there was a deceleration in the output growth of cotton textiles. Based on the relatively early peak of the cotton industry’s expansion, David Greasley and Les Oxley conclude that “it appears unlikely that cotton played the major role in the post-1815 upswing in British industrialization.”49
Following a completely different trajectory, growth in iron goods was more in line with the GPT model. Starting in the 1780s, the growth rate of the British iron industry accelerated to a peak of about 5.3 percent in the 1840s.50 “Compared to cotton textiles, change in iron was gradual, incremental, and spread out over a longer period of time,” Espen Moe writes.51 With a limited role for cotton, the gradual expansion of the iron industry led Britain’s post-1815 industrial surge, as its trend output tracked much more closely with that of aggregate industry. In sum, the cotton industry followed the growth path of a leading sector, whereas developments in the iron industry reflected the impact timeline of a GPT.
The timing of Britain’s mechanization, linked to the expanded uses of iron in machine-making, also aligned with the GPT trajectory. The first metalworking tools for precision engineering, including Wilkinson’s boring mill of 1774 and Maudslay’s all-iron lathe, appeared in the late eighteenth century, but they would remain in a “comparatively rudimentary state” until about 1815.52 According to accounts of qualified engineers and the 1841 Select Committee on Exportation of Machinery, over the course of the next two decades improvements and standardization in such machine tools ushered in a “revolution” in machine-making.53 The gradual evolution of the mechanical engineering industry provides additional support for a delayed impact timeframe for mechanization. According to British patent data from 1780 to 1849, the share of mechanical engineering patents among the total number of patents increased from an average of 18 percent in the decade starting in 1780 to a peak of 34 percent in the one starting in 1830.54
DELAYED, OUT-OF-PERIOD EFFECTS: STEAM ENGINE AND FACTORY SYSTEM
Compared to mechanization, the steam engine had not diffused widely enough through Britain’s economy by the mid-nineteenth century to make a substantial impact on overall industrial productivity. Detailed investigations of steam engine adoption have forced a reassessment of commonly held assumptions about the rapid impact of steam power on British productivity growth.55 According to one growth accounting analysis, which compares the impact of steam against water power as a close substitute source of power, steam power’s contribution to British productivity growth was modest until at least the 1830s and most influential in the second half of the nineteenth century.56 This revised impact timeframe conflicts with international relations scholarship, which advances faster trajectories for steam’s impact as a leading sector.57
Steam engine adoption was slow. In 1800, thirty years after Watt patented his steam engine, there were only about thirty-two engines operating in Manchester, which was a booming center of industrialization.58 Even into the 1870s, many important sectors in the British economy, such as agriculture and services, were virtually unaffected by steam, as most of steam power’s applications were concentrated in mining and in cotton textiles.59 During the period when LS accounts expect its peak growth, the steam engine could not claim generality of use.
The process by which the steam engine gained a broad variety and range of uses entailed complementary innovations that followed many years after the introduction of Watt’s steam engine. It took sixty years for steam to become the prime driver of maritime transport; that was made possible only after cumulative enhancements to the power of steam engines and the replacement of paddle wheels by screw propellers, which increased the speed of steam-powered ships.60 Watt’s original low-pressure design engines consumed large amounts of coal, which hindered widespread adoption. After 1840, aided by inventions such as the Lancashire boiler and discoveries in thermodynamics, it became economically viable to deploy steam engines that could handle higher pressures and temperatures.61 In sum, steam power may be the quintessential example of the long delay between the introduction of a GPT and significant economy-wide effects.
It is worth assessing whether interconnections between developments in steam and those in iron and cotton give grounds for an earlier impact trajectory.62 Yet both forward and backward linkages were limited in the early stages of the steam engine’s evolution. Regarding the latter, the steam engine–producing industry did not substantially stimulate the iron industry’s expansion. In the late 1790s, at a peak in sales, Boulton and Watt steam engines consumed less than 0.25 percent of Britain’s annual iron output.63 Forward linkages to textiles, the most likely sector to benefit from access to cheaper steam power, were also delayed. Steam-powered inventions in textiles did not overtake water-powered textile processes until after 1830.64 Of course, over the long run steam power was a critical technological breakthrough that changed the energy budget of the British economy.65 However, for investigating the mechanisms that facilitated Britain’s rise to become the clear productivity leader in First Industrial Revolution, the steam engine played a modest role and most of its effects were out-of-period.
A similar timeline characterizes the progression of the factory system, another candidate GPT considered for this period. The factory system diffused slowly and took hold in only a limited number of trades during the time when Britain was establishing its industrial preeminence. The British textile industry, as the earliest adopter of this organizational innovation, had established nearly five thousand steam- or water-powered factories by the 1850s.66 Other industries, however, were much slower to adopt the factory system. In the first decades of the nineteenth century, small workshops and domestic production still dominated the metal trades as well as other hardware and engineering trades.67
Moreover, factories were still relatively small even into the mid-nineteenth century, and some industries adopted a mixed factory system in which many processes were outsourced to household workers.68 It was not until steam power overtook water power in the 1830s and 1840s as a source of power in factories that the subsequent redesigns of factory layouts led to large gains in productivity.69
What does this clarified chronology of technological impacts in the IR-1 mean for the explanatory power of the GPT and LS mechanisms? Of the three candidate leading sectors, only cotton textiles, which expanded rapidly and peaked in terms of output growth in the 1780s and 1790s, followed the impact timeframe of a leading sector. As figure 3.2 shows, by 1814 British cotton exports had already surpassed 75 percent of the value they would attain in 1840. Yet Britain sustained productivity growth rates at higher levels than its rivals only in the first decades of the nineteenth century. Thus, the period when cotton should have made its greatest impact on Britain’s industrial ascent does not accord with the timeline of Britain’s industrialization surge.
The hurried timeline of the LS mechanism contrasts with the more delayed impact of other technological advances. As predicted by the GPT mechanism, all three candidate GPTs—mechanization, the steam engine, and the factory system—had little impact on Britain’s industrial rise until after 1815. In fact, the diffusion timelines for the steam engine and factory system were so elongated that their impact on Britain’s rise to industrial preeminence was limited in this period. In 1830, steam engine adoption, as measured by total horsepower installed, was only one-quarter of the 1840 level, whereas by 1830 the level of iron production had reached about 50 percent of its corresponding value in 1840 (see figure 3.2). This is consistent with the steady expansion of mechanization across industry in the early decades of the 1800s as the GPT trajectory most attuned with the timing of Britain’s rise to economic leadership.
figure 3.2 Technological Impact Timeframes in the IR-1. Note: British cotton exports, iron production, and steam engine adoption over time. Source: Robson 1957, 331–35; Mitchell 1988, 280–81; Crafts 2004, 342.
Phase of Relative Advantage: Diffusion of Iron vs. Monopoly Profits from Cotton
Thus far, the empirical evidence has presented a bird’s-eye view of the overall timeline of technological change and industrialization in the IR-1, but there are two other dimensions on which the GPT and LS trajectories diverge. According to the expectations of the LS mechanism, the phase of technological development central to Britain’s relative economic rise was its dominance of key innovations in cotton textiles, iron, or the steam engine. The GPT mechanism predicts, in contrast, that Britain’s advantage in the diffusion of mechanization, the steam engine, or the factory system was the key driver.
The rest of this section tests two sets of predictions derived from the diverging assumptions of the two mechanisms. First, regarding the geographic clustering of major technological breakthroughs in the IR-1, I assess whether innovations in candidate leading sectors and GPTs were concentrated in Britain. Next, regarding the comparative consequences of these technologies, I evaluate whether Britain’s industrial superiority drew more from its advantages in the production and exports of the IR-1’s leading sectors or from its advantage in the diffusion of the IR-1’s GPTs.
INNOVATION CLUSTERING IN THE IR-1’S BREAKTHROUGHS
Did Britain dominate innovation in the leading sectors of the IR-1? At first glance, there is no question that radical advances in candidate leading sectors clustered in Britain. This list includes Watt’s steam engine, Arkwright’s water frame, Cort’s puddling process, and many more. Per one analysis of 160 major innovations introduced during the nineteenth and twentieth centuries, Britain was home to 44 percent of major innovations from 1811 to 1849—a rate that was double that of the closest competitor (the United States at 22 percent).70
Further investigation into British superiority in technological innovation paints a more mixed picture. According to another list of technical advances by country of origin, Britain accounted for only 29 percent of major innovations in the years from 1826 to 1850, a period that corresponds to when it cemented its productivity leadership.71 Moreover, the European continent introduced many significant innovations, including the Jacquard loom, mechanical flax spinning, chlorine bleaching, the Leblanc soda–making process, and the Robert continuous papermaking machine.72 France, in particular, generated many of the major industrial discoveries, such as in chemicals, clocks, glass, papermaking, and textiles.73
Thus, some scholars argue that Britain’s comparative edge was in more incremental improvements. Reflecting on technological creativity in the IR-1, economic historian Joel Mokyr argues, “Britain seems to have no particular advantage in generating macroinventions … the key to British technological success was that it had a comparative advantage in microinventions.”74 A proverb from the time captured this distinction: “For a thing to be perfect it must be invented in France and worked out in England.”75 This suggests that digging deeper into the different phases of technological development can help uncover the roots of Britain’s industrial leadership.
MONOPOLY PROFITS VS. DIFFUSION DEFICIT
First, as was the case with the period when they made their impact, developments in cotton and iron followed very different paths with respect to the phase of technological change that determined economic differentials. Britain’s cotton textile industry, the most likely source of monopoly profits, grew faster than other industries before 1800, and it sold most of its goods abroad. Technological innovations such as the spinning jenny and the water frame triggered exponential increases in the efficiency of cotton production, and Britain’s cotton output increased by 2,200 percent from 1770 to 1815.76 From 1750 to 1801, cotton’s share of Britain’s major exports increased from 1 percent to 39.6 percent.77
Certainly, the growth of British cotton exports was remarkable, but what was the impact of associated monopoly rents on overall growth differentials? Supported by improved quantitative estimates of the cotton industry’s impact on the British economy, historians generally accept that the cotton industry was much more significant for enhancing Britain’s trade balance than for boosting its economic productivity.78 According to one estimate, between 1800 and 1860, the cotton industry accounted for 43 percent of the threefold increase in the value of exports but only 8 percent of the threefold increase in national income.79
Overall, exports constituted a small proportion of British economic activity during the IR-1. From 1770 to 1841, British exports as a percentage of overall industrial demand increased only from 13 to 16 percent.80 Now, these figures probably underrate trade as a critical engine of growth for Britain in the IR-1, as they ignore gains from the reinvestment of profits from overseas trade.81 But the impact of reinvestment has been challenged, and it is not apparent why reinvestments from exports were more important than reinvestments from the profits generated by domestic production.82
The iron industry’s impact on Britain’s economic rise did not run through monopoly profits from innovation. From 1794 to 1796, British ironmakers contributed 11 percent of Britain’s manufacturing exports. This proportion actually declined to just 2 percent by the 1814–1816 period and stayed around that rate into the 1830s.83 It is also questionable whether Britain held a relative advantage in iron exports during the late eighteenth century, which is when LS accounts expect monopoly profits to drive British industrialization.84 In fact, British industries continued to rely on imports of high-grade iron from Sweden and Russia well into the nineteenth century.85
An alternative pathway, captured by the GPT trajectory, posits that Britain’s advantage came from the diffusion of iron machinery advances across a wide range of sectors. To trace this trajectory, it is necessary to pay more attention to what a prominent historian of the IR-1 calls one of the astonishing things about the phenomenon: the gap between “innovation as ‘best practice’ technique and the diffusion of innovation to become ‘representative’ technique.”86
Britain was more successful than its industrial rivals in the diffusion of mechanization. Contemporary observers from the European continent often remarked upon Britain’s ability to bridge this gap between best practice and representative practice.87 Writing in 1786 in their Voyages aux Montagnes, French observers F. and A. de la Rochefoucauld-Liancourt, commenting on Britain’s relative advantage in the widespread adaptation of the use of iron, noted
the great advantage [their skill in working iron] gives them as regards the motion, lastingness and accuracy of machinery. All driving wheels and in fact almost all things are made of cast iron, of such a fine and hard quality that when rubbed up it polishes just like steel. There is no doubt but that the working of iron is one of the most essential of trades and the one in which we are most deficient.88
But France’s deficiency in iron machinery was not a product of its lack of access to key innovations. In fact, France was the world’s center of science from the late eighteenth century until the 1830s.89 Rather, as the following quote illustrates, Britain’s industrial rivals fell behind in “diffused average technology” and the “effective spread of technical change more widely.” Economic historian Peter Mathias writes:
It is remarkable how quickly formal knowledge of “dramatic” instances of new technology, in particular steam engines, was diffused, and how quickly individual examples of “best-practice” technology in “show piece” innovations were exported. The blockage lay in the effective spread of technical change more widely—diffused average technology rather than single instances of best-practice technology in “dramatic” well-publicized machines.90
Advances in iron metallurgy played a crucial role in a GPT trajectory that spread from a sector that improved the efficiency of producing capital goods. The GPT trajectory unfolds as the technology becomes more general-purpose through interactions between the upstream capital goods sector and the user industries that enlarge the range of applications. Rosenberg’s depiction of this type of system highlights the nineteenth-century American machine tool industry as the innovative capital goods sector.91 In this case, Britain’s metal-processing works were the crucial wellspring. Specifically, technical advances in iron fed into metalworking industries from which broadly similar production processes diffused over a large number of industries.92 Maxine Berg, a professor of history at the University of Warwick, pinpoints these industries as the “prime mechanism for technological diffusion.”93
Scholars also identify Watt’s improved steam engine as a potential source of both LS- and GPT-based effects. Here I focus on testing the LS prediction about the steam engine–producing industry because the previous section showed that the steam engine and the factory system, as candidate GPTs, diffused too slowly to make a meaningful impact on the economic power transition in the IR-1.
It is tough to make a case that the growth of the steam engine–producing industry generated a substantial source of monopoly profits for Britain. Equipped with an exclusive patent, James Watt and Matthew Boulton set up a firm in 1775 to sell steam engines.94 In the period from 1775 to 1825, however, the firm sold only 110 steam engines to overseas customers.95 By 1825, France and the United States were manufacturing the Watt engine and high-pressure engines at more competitive prices, and overseas demand declined sharply.96 Thus, the international sales history of this firm severely weakens the significance of the monopoly profits associated with the innovation of the steam engine.97
In sum, the evidence from this section supports two conclusions. British advantages in the production and export of iron, steam engines, and cotton textiles (the best representative of the LS trajectory) had muted effects on its overall industrialization and productivity advances. Second, the contributions of technological breakthroughs in iron metallurgy and steam power to Britain’s industrial rise track better with the GPT mechanism, based on relative advantages in widespread technological diffusion as opposed to monopoly profits from innovation.
Breadth of Growth: Complementarities of Iron vs. Spillovers from Cotton Textiles
The breadth of growth in the IR-1 is the last dimension on which the LS and GPT trajectories disagree. Was Britain’s industrial rise driven by technological changes confined to a narrow range of leading sectors, or was it based on extensive, complementary innovations that enabled the spread of GPTs? Making use of data on sectoral sources of productivity growth, trade flows, and patents, I evaluate these competing propositions about the breadth of technological change in the industrial revolution.
WIDESPREAD PRODUCTIVITY GROWTH
In differentiating between the narrow view and the broad view of technical change during the IR-1, a natural starting point is to estimate the contribution of various industries to British productivity growth. Deirdre McCloskey’s calculations of sectoral contributions to productivity growth support the broad view. Though cotton accounted for a remarkable 15 percent of Britain’s total productivity growth, nonmodernized sectors still drove the lion’s share (56 percent) of productivity gains.98
Manufacturing trade data provide another testing ground. If other manufacturing industries outside of textiles and iron were technologically stagnant during the first fifty years of the nineteenth century, then British competitiveness in these industries should decline relative to textiles and iron. The narrow view implies that Britain should have imported other manufactures. Peter Temin’s analysis of British trade data, however, finds the opposite. Throughout the first half of the nineteenth century, British manufacturing exports matched the increase in cotton exports throughout the first half of the nineteenth century.99 In a wide range of manufactures, such as arms and ammunition, carriages, glass, and machinery and metals, Britain held a clear comparative advantage. This pattern points to some general pattern of change that spanned industries. “The spirit that motivated cotton manufactures extended also to activities as varied as hardware and haberdashery, arms, and apparel,” Temin concludes.100
The patent record also depicts a landscape of extensive technological change.101 From 1780 to 1840, about 80 percent of all patented inventions came from outside the textiles and metals industries.102 Per Christine MacLeod’s data on British patents covering the 1750–1799 period, most capital goods patents originated from sectors outside of textile machinery and power sources.103 As summed up by historian Kristine Bruland, the historical evidence supports “the empirical fact that this was an economy with extensive technological change, change that was not confined to leading sectors or highly visible areas of activity.”104
GPTS AND COMPLEMENTARY INNOVATIONS
At this point, indicators of the multisectoral spread of innovation in the IR-1 should not be sufficient to convince a skeptical reader of the GPT mechanism’s validity. Broad-based growth could be a product of macroeconomic factors, such as sound fiscal and monetary policy or labor market reforms, rather than a GPT trajectory.105 Proving that the dispersion of technological change in Britain’s economy reflected a GPT at work requires evidence that connects this broad front to mechanization.106
Input-output analysis, which sheds light on the linkages between industries, suggests that improvements in the working of iron had broader economic significance. To better understand the interrelationships among industries during the industrial revolution, Sara Horrell, Jane Humphries, and Martin Weale constructed an input-output table for the British economy in 1841. Across the seventeen industries included in the analysis, the two industries most closely associated with mechanization—metal manufacture and metal goods—scored the highest on combined backward and forward linkages.107 These two domains were “lynchpins of linkage effects.”108
Patent indicators confirm these results. When patents are grouped according to standard industry taxonomies, the resulting distribution shows that the textile industry contributed to 15 percent of the patents issued between 1711 and 1850, making it the most inventive industry in aggregate terms.109 However, when patents are sorted by general techniques as opposed to industry sectors, the same data reveal the underlying drive force of mechanical technology: it is linked to almost 50 percent of all British patents during this period.110
Along all three dimensions of technological trajectories in the IR-1, the process-tracing evidence bolsters the validity of the GPT mechanism. First, slower-moving developments in mechanization lined up with a delayed timeline of Britain’s industrialization. Other candidate leading sectors and GPTs either peaked too early (cotton) or got started too late (steam engine, factory system). Second, Britain gained its industrial dominance from a relative advantage in widespread adoption of iron metalworking and linked machinery. Third, the benefits from this GPT advantage circulated throughout the economy, rather than remaining concentrated in the iron industry.
The standard explanation of how the IR-1 gave rise to a power transition, as captured by the LS mechanism, analyzes technological change at the level of industries that grow faster than others. The historical evidence reveals the limitations of these industry taxonomies. Instead, advantages in the diffusion of production machinery—a general pattern of change that extended across a wide range of economic activities—propelled Britain to industrial dominance.111
Institutional Complementarities: GPT Skill Infrastructure in the IR-1
Having mapped Britain’s industrial rise to a GPT trajectory linked to mechanization, there is still a need to explain why Britain was best positioned to exploit this trajectory. If other countries at the technological frontier can also cultivate mechanical innovations at home and absorb them from abroad, why were Britain’s competitors unable to benefit from the diffusion of metalworking processes to the same extent? This section supports an explanation based on Britain’s institutional competencies in widening the pool of engineering skills and knowledge linked to mechanization.
Which types of institutions for skill provision were most conducive to national success in the IR-1? One common refrain is that Britain’s leadership was rooted in the genius of individual innovators like James Watt, and such genius did not transfer as quickly across borders during the IR-1.112 Though recent scholarship has weakened this view, many influential histories center on the “heroic” inventors of the industrial revolution.113 Consistent with the LS template, this view focuses on the institutions that helped drive heroic invention in Britain, such as the development of a patent system.114
The pathway by which mechanization propelled Britain’s industrial ascent, established as a GPT trajectory in the previous section, emphasizes another set of institutions for skill formation. In line with GPT diffusion theory, Britain owed its relative success in the IR-1 to mechanics, instrument-makers, and engineers who could build machines according to blueprints and improve upon them depending on the application context. Under this view, the institutions that trained the “tweakers” and “implementers,” rather than those that cultivated genius inventors, take center stage.115
Widening the Base of Mechanical “Tweakers” and “Implementers”
At first, rapid advances in precise metalworking exposed a skills shortage in applied mechanics. Beginning in the 1770s, a cascade of recruitment advertisements in local newspapers sought out an “engine-maker” or a “machine-maker.”116 Reflecting on this skills mismatch, the president of Britain’s Institute of Civil Engineers stated that the use of cast iron in machine parts “called for more workmen than the millwright class could supply.”117
A number of institutional adjustments helped Britain meet this demand for mechanically skilled tweakers and implementers. Initially, Britain benefited from a flexible apprenticeship system that empowered workers in related domains to get trained in applied mechanics.118 Thus, to develop the workforce to build and maintain the machinery of the IR-1, Britain could draw from a wide pool of blacksmiths, millwrights, gunsmiths and locksmiths, instrument-makers, mechanics, and toolmakers.119
In addition, institutes dedicated to broadening the base of mechanical expertise helped diffuse ironmaking and machine-making skills. Starting in the 1790s, private and informal initiatives created a flurry of trade associations that supported a new class of mechanical and civil engineers and helped connect them with scientific societies and entrepreneurs.120 Critical centers included the Andersonian Institution in Glasgow, the Manchester College of Arts and Sciences, the School of Arts in Edinburgh, the Mechanical Institution in London, the Society for the Diffusion of Useful Knowledge, and hundreds of mechanics’ institutes.121 These institutes helped to absorb knowledge from foreign publications on science and engineering, recruit and upskill new mechanical engineers from a variety of trades, and spread mechanical engineering knowledge more widely.122
It is important to note that these institutional features differed from those more suited to the “heroic inventor” model. In Britain’s cotton textile industry, the classic leading sector of the IR-1, the key institutional complements deviated greatly from the education and training systems that widened the base of mechanical expertise in the IR-1. Collating information from biographies of British engineers, online databases, and detailed economic histories, Ralf Meisenzahl and Joel Mokyr constructed a database of 759 British individuals who made incremental improvements to existing inventions during the industrial revolution.123 Notably, based on their analysis of interactions between tweakers and their institutional surroundings, they found that the textile industry was an outlier in terms of protectiveness over intellectual property rights and reluctance to share information about new techniques. Less than one-tenth of tweakers in textiles published their knowledge to a broader audience or joined professional societies, in stark contrast to the two-thirds of tweakers in mechanically inclined fields who did so.124 Over 80 percent of the tweakers who were active primarily in textiles took out at least one patent, compared to just 60 percent for tweakers overall.125
These trends in applied mechanics underscore the significance for British mechanization of “collective invention,” a process that involved firms sharing information freely with one another and engineers publishing technical procedures in journals to spur the rapid diffusion of best-practice techniques. According to one analysis of how various districts adapted to the early phases of industrialization, areas that practiced collective invention often cultivated “a much higher degree of technological dynamism than locations which relied extensively on the patent system.”126
Britain’s Comparative Advantage over France and the Netherlands in GPT Skill Infrastructure
Britain’s competitors also grasped the significance of Britain’s wide pool of mechanical skills. Whereas codified knowledge crisscrossed western Europe and North America via patent specifications, global exchanges among scientific societies, and extensive visits by foreign observers to British workshops and industrial plants, the European continent struggled greatly to absorb tacit knowledge, especially the know-how embodied in the practical engineering skills of British mechanical tweakers and implementers.127 France and the Netherlands fiercely poached British engineers, as the transfer of tacit knowledge in the fields of large-scale ironworking and machine construction almost always necessitated the migration of skilled workers from Britain.128 “It was exactly in the skills associated with the strategic new industries of iron and engineering that [Britain’s] lead over other countries was most marked,” argues Mathias.129
Why did this repository of engineering skills develop more fruitfully in Britain than in its industrial rivals? A growing body of evidence suggests that Britain’s institutions better adapted its distribution of skills to mechanization. Britain’s institutional advantage was rooted in the system of knowledge diffusion that connected engineers with entrepreneurs, cities with the countryside, and one social class with another. Institutes that trained mechanics took part in a broader “mushrooming of associations” that spread technical knowledge in early-nineteenth-century Britain.130 By the mid-nineteenth century, there were 1,020 such associations in Britain, with a total membership of approximately 200,000; clearly, these networks are essential to any explanation that links human capital to Britain’s industrial ascent.131 Compared to their peers on the continent, British mechanics had superior access to scientific and technical publications.132 As a result, the British system of the early nineteenth century had no match in its abundance of people with “technical literacy.”133
The French system, by way of comparison, lacked similar linkages and collaborations between highly educated engineers and local entrepreneurs.134 Though France produced elite engineers at schools like the École Polytechnique, it trained too few practitioners to widen the base of mechanical skills.135 For example, Napoleon’s early-nineteenth-century reform of France’s higher education system encouraged the training of experts for narrow political and military ends, thereby limiting the ability of trainees to build connections with industry.136 These reforms and other industrial policies directed French engineers toward projects associated with luxury industries and specialized military purposes, which “tended to become locked away from the rest of the economy in special enclaves of high cost.”137 To illustrate, through the mid-1830s, only one-third of École Polytechnique graduates entered the private sector.138 France’s system for disseminating mechanical knowledge and skills was vastly inferior to that of the British.
The Netherlands also failed to develop a base of mechanical skills that linked scientific research to practical ends. In some mechanical sciences, the Dutch generated plenty of potentially useful innovations, even pioneering key breakthroughs that eventually improved the steam engine.139 Yet the Dutch struggled to translate these scientific achievements into practical engineering knowledge because they trailed the British in forming institutional settings that made widespread knowledge of applied mechanics possible. Records of Dutch educational systems, the dearth of societies that held lectures and demonstrations for mechanical learning, and the materials available at libraries in technical colleges all “reflected a profound lack of interest in applied mechanics.”140 In his study of Dutch technological leadership, Karel Davids argues that, during the first three-quarters of the eighteenth century, “collaboration between science and industry in the Netherlands failed to merge in the very period that relations between the two became rapidly closer in Britain.”141
Britain’s advantage in GPT diffusion was not rooted in its higher education system, which lagged far behind the French education system during the IR-1 period.142 France had already established more than twenty universities before the French Revolution. The French system of higher technical education, from the late eighteenth century through the 1830s, had no rival. The Grande Écoles system, including the elite École Polytechnique (established in 1794), trained expert scientists and engineers to take on top-level positions as industrial managers and high-level political personnel.143 Up until 1826, England had set up only two universities, Oxford and Cambridge. These institutions made limited contributions to training the workforce necessary for industrialization. One study with a sample of 498 British applied scientists and engineers born between 1700 and 1850 found that only 50 were educated at Oxford or Cambridge; 329 were not university-educated.144
At this point, curiosity naturally leads us to ask why Britain accumulated an advantage in GPT skill infrastructure. Due to practical constraints of time and space, I acknowledge but do not delve into the deeper causes for the notable responsiveness of Britain’s institutions to the skill demands of mechanization. In surveying valuable lines of inquiry on this subject, chapter 2 points to government capacity to adopt long time horizons and reach intertemporal bargains. In the IR-1 case, two specific factors are also worthy of consideration. Attributing Britain’s later success to pre-industrial training practices, some studies suggest that Britain’s apprenticeship system allowed for agile and flexible adaptation to fluctuations in the demand for skills, especially in mechanical trades.145 Looking even further back, other scholars probe the geographical origins of Britain’s mechanical skills, underscoring the lasting effects of Britain’s adoption of watermills in the early Middle Ages.146
Alternative Explanations of Britain’s Rise
The history of the IR-1 is certainly not a neglected topic, and the literature features enthusiastic debates over a wide range of possible causes for Britain’s rise. Prominent explanations tie Britain’s early industrialization to population growth,147 demand and consumption standards,148 access to raw materials from the colonies,149 slavery,150 and trade.151 The obvious concern is that various contextual factors may confound the analysis of the LS and GPT mechanisms.
I am not rewriting the history of the IR-1. I am drawing from one particularly influential and widely held view of the IR-1—that technological advances drove Britain’s industrial ascent—and investigating how technological change and institutional adaptations produced this outcome. The most relevant alternative factors, therefore, are those that provide a different interpretation of how technologies and institutions coevolved to result in Britain’s industrial hegemony. Although I primarily focus on the LS mechanism as the most formidable alternative explanation to the GPT diffusion theory, other explanations also warrant further investigation.
Threat-Based Explanations
Threat-based theories assert that external threats are necessary to incentivize states to innovate and diffuse new technologies. Did Britain owe its technological leadership to war and its military’s impetus to modernize? During the IR-1 period, Britain was embroiled in the Revolutionary and Napoleonic Wars (1793–1815), a near-continuous stretch of conflicts involving France and other European states. If threat-based explanations stand up in the IR-1 case, then the historical record should show that these wars made an essential and positive contribution to Britain’s adoption of iron machinery and mechanization.
Some evidence supports this argument. By 1805, the British government’s needs for iron in the war effort accounted for 17 percent of the total British iron output in 1805.152 This wartime stimulus to iron production facilitated improvements in iron railways, iron ships, and steam engines.153 In particular, military investments in gunmaking produced important spin-offs in textiles and machine tools, most famously encapsulated by Watt’s dependence on John Wilkinson’s cannon boring techniques to make the condenser cylinders for his steam engine.154
On the flip side, war’s disruptive costs are likely to have offset any stimulus to Britain’s mechanization. Aside from Wilkinson’s cannon boring device and some incremental improvements, wartime pressures did not produce any major technological breakthroughs for the civilian economy.155 Military needs absorbed productive laborers from Britain’s civilian economy, resulting in labor shortages.156 War also limited both the domestic demand for iron, by halting investment in construction, agriculture, and other industries, and the foreign demand for British iron, by cutting off foreign trade. Historian Charles Hyde notes, “In the absence of fighting, overall demand for iron might have been higher than it was.”157
Furthermore, any temporary benefits that accrued to Britain’s iron industry in wartime were wiped out in the transition to peacetime. In one influential text, historian Thomas Ashton reviewed how each of the wars of the eighteenth century, including the Revolutionary and Napoleonic Wars, affected Britain’s iron industry.158 He observed a similar pattern in each case. At first, the outbreak of hostilities boosts demand for iron in the form of armament, and trade disruptions protect domestic producers against foreign competitors. This initial boom is followed by a severe crash, however, when the iron industry adjusts to the conflict’s aftermath. A trade depression follows. Converting foundries to make plowshares instead of cannons incurs heavy losses, made even more painful by the fact that war conditions promoted “feverish” developments that were unsustainable in the long run.159
On a more fundamental level, threat-based theories have limited leverage in explaining Britain’s relative rise because its economic competitors were also embroiled in conflicts—in many cases, against Britain. The Dutch fought Britain in the fourth Anglo-Dutch War (1780–1784) as well as in the Napoleonic Wars.160 Of course, during this time, France was Britain’s main military opponent. Thus, since the Netherlands and France also faced a threatening external environment, the net effect of the war on economic growth differentials should have been minimal.161 If anything, since France fought on many more fronts than Britain during this period, proponents of threat-based explanations would expect France to have experienced more effective and widespread diffusion of iron machinery throughout its economy. The case analysis clearly discredits that expected outcome.
VoC Explanations
Can Britain’s particular brand of capitalism account for its technological rise? The varieties of capitalism (VoC) approach posits that liberal market economies (LMEs) are particularly suited to radical innovation. Consistent with this framework, international political economy scholars emphasize that Britain’s free market economy supported gains in rapidly changing technological domains like consumer goods, light machine tools, and textiles.162 During the IR-1 period, Britain began to develop the institutional features that would cement it as a LME, including decentralized collective bargaining and high levels of corporatization.163 Most pertinent to GPT diffusion theory, VoC scholars expect LMEs like Britain to excel at cultivating general skills, which help transfer GPT-related knowledge and techniques across firms.
Taking measure of Britain’s human capital development in general skills in this period is therefore central to evaluating whether its technological leadership can be explained by its form of capitalism. Overall, estimates of literacy rates and school attendance demonstrate that the general level of human capital in Britain was notably low for an industrial leader.164 British literacy rates for males were relatively stagnant between 1750 and 1850, and average literacy rates in Britain were much lower than rates in the Netherlands and barely higher than those in France around the turn of the nineteenth century.165 In fact, general levels of educational attainment in Britain, as measured by average years of schooling, declined from around 1.4 years in 1740 to 1.25 years in 1820.166 Contrary to VoC theory’s expectations, Britain did not hold an advantage in general skills during this period.
The VoC explanation’s applicability to the IR-1 period is further limited by issues with designating Britain as the only LME in this period. Like Britain, the Netherlands functioned as a relatively open economy and exhibited tendencies toward economic liberalism, but it was not able to adapt and diffuse significant technological changes.167 Though France is now considered a coordinated market economy, in the early nineteenth century it took on some of the characteristics of LMEs by implementing capital market reforms and trade liberalization.168 The VoC approach therefore struggles to resolve why these two LMEs diverged so greatly in their adaptation to mechanization.
Case-Specific Factors
Among other factors specific to the IR-1 setting, one alternative explanation emphasizes Britain’s fortunate geographic circumstances. More specifically, classic works have argued that proximity to plentiful coalfields was essential to British industrialization.169 These natural resource endowments enabled the expansion of coal-intensive industries, such as the iron industry. In this line of thinking, the fact that coal was cheaper in Britain than elsewhere in Europe explains why Britain was the first to sustain productivity leadership.170
The relationship between coal and industrialization does not necessarily undermine the GPT mechanism. For one, in principle, Britain’s competitors could also have effectively leveraged coal resources. The southern provinces of the Netherlands were located close to Belgian coalfields.171 Over the course of the eighteenth century, Dutch industry had mostly shifted to coal, and away from peat stocks, as a key source of energy.172 Even if Britain’s industrial rivals had to pay more by importing coal, the expected productivity gains associated with adopting new technologies should have outweighed these costs. Moreover, GPT skill infrastructure could have mediated the relationship between coal and mechanization, as Britain’s edge in metalworking skills spurred the adoption of new coal-using technologies, which strengthened the connection between proximity to coal and economic growth.173
Summary
In many ways, the industrial revolution marked an exceptional transformation. It is to any number of historical trends what the birth of Jesus is to the Gregorian calendar—an inflection point that separates “before” and “after.” For my purposes, however, the industrial revolution is a typical case showing how technological revolutions influence the rise and fall of great powers. Evidence from great powers’ different adaptations to technological changes in this period therefore helps test GPT diffusion theory against the LS mechanism.
In sum, GPT diffusion theory best explains why Britain led Europe’s industrial transformation in this period. Britain effectively capitalized on general-purpose improvements in mechanization owing to its institutional advantages that were conducive to widening the pool of mechanical skills and knowledge. According to GPT diffusion theory, countries like this disproportionately benefit from technological revolutions because they adapt more successfully to the GPT trajectories that transform productivity. In line with these expectations, Britain was more successful than its industrial rivals in sustaining long-term economic growth, which became the foundation of its unrivaled power in the early and mid-nineteenth century.
On the flip side, this chapter’s case analysis undercuts the LS-based explanation. The timeframe for when leading sectors were expected to stimulate Britain’s productivity growth did not align with when Britain’s industrialization took off. Britain’s economic ascent owed more to the widespread adoption of iron metalworking and linked production machinery than to monopoly profits from cotton textiles. The key institutional complements were not those that produced heroic inventions—Britain’s rivals held their own in these areas—but rather those that fostered widespread knowledge of applied mechanics.
Do these findings hold in other periods of technological and geopolitical upheaval? The IR-1 was one of the most extraordinary phases in history, but it was not the only era to attain the title of an “industrial revolution.” To further explore these dynamics, it is only appropriate to turn to the period some have labeled the Second Industrial Revolution.
4 The Second Industrial Revolution and America’s Ascent
IN THE LATE nineteenth and early twentieth centuries, the technological and geopolitical landscape transformed in ways familiar to observers of today’s environment. “AI is the new electricity” goes a common refrain that compares current advances in machine intelligence to electrical innovations 150 years ago. Those fundamental breakthroughs, alongside others in steel, chemicals, and machine tools, sparked the Second Industrial Revolution (IR-2), which unfolded from 1870 to 1914.1 Studies of how present-day technological advances could change the balance of power draw on geopolitical competition for technological leadership in the IR-2 as a key reference point.2
Often overshadowed by its predecessor, the IR-2 is equally important for investigating causal patterns that connect technological revolutions and economic power transitions. The presence of both cause and outcome ensures a fruitful test of the GPT and LS mechanisms. The beginning of the period featured remarkable technological innovations, including the universal milling machine, the electric dynamo, the synthesis of indigo dye, and the internal combustion engine. According to some scholars, one would be hard-pressed to find another period with a higher density of important scientific advances.3 By the end of the period, Britain’s decline and the rise of Germany and the United States had yielded a new balance of economic power, which one historian describes as a “shift from monarchy to oligarchy, from a one-nation to a multi-nation industrial system.”4 Arguably, British industrial decline in the IR-2 was the ultimate cause of World War I.5
International relations scholars hold up the IR-2 as a classic case of a power transition caused by LS product cycles.6 According to this view, Britain’s rivals cornered market shares in the new, fast-growing industries arising from major technological innovations in electricity, chemicals, and steel.7 Specifically, scholars argue that Germany surpassed Britain in the IR-2 because it was “the first to introduce the most important innovations” in these key sectors.8 Analysis of emerging technologies and today’s rising powers follows a similar template when it compares China’s scientific and technological capabilities to Germany’s ability to develop major innovations in chemicals.9 Thus, as a most likely case for the LS mechanism, which is favored by background conditions and existing theoretical explanations, the IR-2 acts as a good test for the GPT mechanism.
Historical evidence from this period challenges this conventional narrative. No country monopolized innovation in leading sectors such as chemicals, electricity, steel, and motor vehicles. Productivity growth in the United States, which overtook Britain in productivity leadership during the IR-2, was not dominated by a few R&D-based sectors. Moreover, major breakthroughs in electricity and chemicals, prominent factors in LS accounts, required a protracted process of diffusion across many sectors before their impact was felt. This made them unlikely key drivers of the economic rise of the United States before 1914.
Instead, the IR-2 case evidence supports GPT diffusion theory. Spurred by inventions in machine tools, the industrial production of interchangeable parts, known as the “American system of manufacturing,” embodied the key GPT trajectory.10 The United States did not lead the world in producing the most advanced machinery; rather, it had an advantage over Britain in adapting machine tools across almost all branches of industry. Though the American system’s diffusion also required a long gestation period, the timing matches America’s industrial rise. Incubated by the growing specialization of machine tools in the mid-nineteenth century, the application of interchangeable parts across a broad range of manufacturing industries was the key driving force of America’s relative economic success in the IR-2.11
Since a nation’s efficacy in adapting to technological revolutions is determined by how well its institutions complement the demands of emerging technologies, the GPT model of the IR-2 highlights institutional factors that differ from those featured in standard accounts. LS-based theories tend to highlight Germany’s institutional competencies in scientific education and industrial R&D.12 In contrast, the case analysis points toward the American ability to develop a broad base of mechanical engineering skills and standardize best practices in mechanical engineering. Practice-oriented technical education at American land-grant colleges and technical institutes enabled the United States to take better advantage of interchangeable manufacturing methods than its rivals.
This chapter’s evidence comes from a variety of sources. In tracing the contours of technological trajectories and the economic power transition in this period, I relied on histories of technology, categorization schemes from the long-cycle literature, general accounts of economic historians, and revised versions of historical productivity measures. I investigated the fit between institutions and technology in leading economies using annual reports of the US Commissioner of Education, British diplomatic and consular reports, cross-national data on technological diffusion, German engineering periodicals, and firsthand accounts from inspection teams commissioned to study related issues.13 My analysis benefited from archival materials based at the Bodleian Library’s Marconi Archives (United Kingdom), the Library of Congress (United States), and the University of Leipzig and from records of the British Foreign Office.
This chapter proceeds as follows. I begin by chronicling the economic power transition that took place during the IR-2 to clarify that the United States, not Germany, ascended to industrial preeminence. I then identify the key technological breakthroughs, which I sort according to their ties to GPT and LS trajectories. Along the dimensions of impact timeframe, phase of relative advantage, and breadth of growth, this chapter demonstrates that the GPT trajectory aligns better with how the IR-2 enabled the economic rise of the United States. Next, I evaluate whether differences in GPT skill infrastructure can account for the American edge over Britain and Germany in interchangeable manufacturing methods. Toward the chapter’s end, I also tackle alternative explanations.14
A Power Transition: America’s Ascent
To begin, tracing when an economic power transition takes place is critical. In 1860, Britain was still at the apogee of its industrial power.15 Most historians agree that British industrial preeminence eroded in the late nineteenth century. By 1913, both the United States and Germany had emerged as formidable rivals to Britain with respect to the industrial and productive foundations of national power. According to Paul Kennedy’s influential account, before World War I Britain was “in third place,” and “in terms of industrial muscle, both the United States and imperial Germany had moved ahead.”16 Aided with more data than was available for the IR-1 case, I map the timeline of this economic power transition with various measures of industrial output and efficiency.
In the IR-2 case, clarifying who surpassed Britain in economic efficiency takes on added gravity. Whereas in the IR-1 Britain separated itself from the rest, both the United States and Germany challenged British industrial power in the IR-2. But studies of this case often neglect the rise of the United States. Preoccupied with debates over whether Germany’s overtaking of Britain sparked World War I, the power transition literature has directed most of its attention to the Anglo-German competition for economic leadership.17 Some LS-based accounts explain only Germany’s rise in this period without investigating America’s ascent.18
As the rest of this section will show, Germany and the United States both surpassed Britain on some measures of economic power, but the United States emerged as the clear productivity leader. Therefore, any explanation of the rise and fall of technological leadership in this period must be centered on the US experience. The following sections trace the contours of the IR-2’s economic power transition with a range of indicators for productivity leadership, including GDP per capita, industrialization, labor productivity, and total factor productivity.
GDP PER-CAPITA INDICATORS
Changes in total GDP over the course of the IR-2 provide a useful departure point for understanding changes in the balance of productive power. At the beginning of the period in 1871, Germany’s economy was around three-quarters the size of the British economy; by the end of the period, in 1913, Germany’s economy was approximately 14 percent larger than Britain’s. The growth trajectory of the American economy was even starker. Over the same time period, overall economic output in the United States increased from 1.2 times to around 3.4 times that of Britain’s total GDP.19 This trend is further confirmed by the growth rates of overall GDP for the three countries. In the period between 1870 and 1913, the US GDP grew roughly 5.3 times over, compared to 3.3 for Germany and 2.2 for the United Kingdom.20
While gross economic size puts countries in contention for economic leadership, the most crucial outcome is sustained economic efficiency. Compared to total output, trend lines in real GDP per capita mark out a broadly similar picture of the IR-2 period, but they also differ in two significant respects (figure 4.1). First, whereas the United States was already the largest economy by total output in 1870, the United Kingdom maintained a slight lead in real GDP per capita over the United States in the 1870s. The United Kingdom’s average GDP per capita over the decade was about 15 percent higher than the US equivalent.21 US GDP per capita was roughly on par with Britain’s throughout the 1880s and 1890s, but the United States established a substantial lead starting around 1900.22
FIGURE 4.1. Economic Power Transition during the IR-2. Source: Maddison Project Database, version 2020 (Bolt and van Zanden 2020).
Second, in contrast to trend lines in aggregate economic output, Germany did not surpass Britain in GDP per capita before World War I. Germany certainly closed the gap, as its GDP per capita increased from around 50 percent of British GDP per capita in 1870 to around 70 percent in the years before World War I. However, Germany never even came close to overtaking the United Kingdom in GDP per capita during this period.23 This is an important distinction that justifies the focus on US technological success in the IR-2, since surpassing at the technological frontier is a different challenge than merely catching up to the technological frontier.
INDUSTRIALIZATION INDICATORS
Industrialization indicators back up the findings from the GDP per-capita data. The United States emerged as the preeminent industrial power, boasting an aggregate industrial output in 1913 that equaled 36 percent of the global total—a figure that exceeded the combined share of both Great Britain and Germany.24 More importantly, the United States became the leading country in terms of industrial efficiency, with a per-capita industrialization level about 10 percent higher than Britain’s in 1913.25
Once again, the emphasis on productivity over aggregate output reveals that the economic gap between Germany and Britain narrowed but did not disappear. In aggregate terms, Germany’s share of the world’s industrial production rose to 16 percent in 1913. This eclipsed Britain’s share, which declined from 32 percent of the world’s industrial production in 1870 to just 15 percent in 1913.26 However, Germany did not overtake Britain in industrial efficiency. In 1913, its per-capita industrialization level was about 75 percent of Britain’s.27 The magnitude of this gap was approximately the same as the gap between German per-capita GDP and British per-capita GDP.
PRODUCTIVITY INDICATORS
Lastly, I consider various productivity statistics. Stephen Broadberry’s work on the “productivity race” contains the most comprehensive and rigorous assessments of productivity levels in Britain, Germany, and the United States in this period.28 Comparative statistics on labor productivity line up with findings from other indicators (figure 4.2). The United States surpassed Britain in aggregate labor productivity during the 1890s or 1900s, whereas Germany’s aggregate labor productivity increased relative to but did not fully overtake Britain’s over the IR-2 period.29
FIGURE 4.2. Comparative Labor Productivity Levels in the IR-2. Source: Broadberry 2006, 110.
Another set of productivity indicators, Maddison’s well-known and oft-cited historical data on comparative GDP per hour worked, supports Broadberry’s comparative measures of labor productivity levels.30 According to Maddison’s estimates of the average rate of productivity growth from 1870 to 1913, the American and German economies were both growing more productive relative to the British economy. The growth rate of America’s GDP per hour worked was 1.9 percent compared to 1.8 percent for the German rate and 1.2 percent for the UK rate.31
It should be noted that the United Kingdom may have retained a total factor productivity lead in this period. Based on 1909 figures, the last measurements available before World War I, the US aggregate TFP was a little over 90 percent of Britain’s. By 1919, US aggregate TFP was nearly 10 percent larger than Britain’s.32 The United States could have surpassed Britain in overall TFP before World War I, but the data do not clearly demonstrate this outcome. Still, the TFP data track well with the general trends found in other measures of economic efficiency, including a marked increase in US TFP in the 1890s and 1900s as well as a steady narrowing of the gap between UK and German TFP throughout the period. Issues related to the availability, reliability, and comparability of capital stock estimates during this period, however, caution against concluding too much from the TFP trends alone.33
Albeit with some caveats, the general thrust of evidence confirms that the United States overtook Britain in productivity leadership around the turn of the twentieth century. In productive efficiency, Germany significantly narrowed the gap but did not surpass Britain. A clarified picture of the outcome also helps guide the assessment of the LS and GPT mechanisms. In contrast to work that focuses on Anglo-German rivalry in this period, I prioritize explaining why the United States became the preeminent economic power. Moreover, if GPT diffusion theory holds for this period, it should also explain why the United States was more successful than Germany in overtaking Britain in productivity during this period.
Key Technological Changes in the IR-2
Which technological changes could have sparked the economic power transition before World War I? The IR-2 was an age of dizzying technological breakthroughs, including but not limited to the electric dynamo (1871), the first internal combustion engine (1876), the Thomas process for steel manufacturing (1877), and the synthesis of indigo dye (1880).34 Tracking down how every single technical advance could have affected the growth differentials among Britain, Germany, and the United States is an unmanageable task. I narrow the scope of analysis to the most likely sources of LS and GPT trajectories based on previous scholarship that calls attention to the significance of certain technological developments in the IR-2. Once confirmed to meet the established criteria for leading sectors and GPTs, these technological drivers serve as the fields of reference for assessing the validity of the GPT and LS mechanisms in this case.
Candidate Leading Sectors
I focus on the chemicals, electrical equipment, motor vehicles, and steel industries as the leading sectors of the IR-2. These choices are informed by scholars who study the implications of technological change during this period from a LS perspective. The first three sectors feature in the standard rendering of the IR-2 by prominent historical accounts, which centers major discoveries in chemistry and electricity as well as the invention of the internal combustion engine.35 Among those who study the effect of technological revolutions on the balance of power, there is near-consensus that the chemicals and electrical industries were technologically advanced, fast-growing industries during this time.36 Some scholars also identify the automobile industry as a key industry in this period.37 Others reason, however, that automobiles did not emerge as a leading sector until a later period.38
The automobile, chemicals, and electrical industries all experienced prodigious growth during the IR-2, meeting the primary qualification for leading sectors. According to statistics from the US census, the percentage increase in value added by manufacture in each of the chemicals, electrical, and automobile industries was much higher than the average across all industries from 1899 through 1909. In fact, the automobile and electrical equipment industries boasted the two highest rates of percentage growth in value added over this period among sectors with a market size over $100 million.39
I also consider developments in steel as a possible source of leading-sector product cycles. It is hard to ignore the explosive growth of the steel industry in both Germany, where it multiplied over 100-fold from 1870 to 1913, and the United States, where it multiplied around 450 times over the same period.40 In addition, many scholars list steel as one of the leading sectors that affected the economic power balance in the IR-2.41 Rostow identifies steel as part of “the classic sequence” of “great leading sectors.”42 In sum, I consider four candidate leading sectors in this period: the automobile, chemicals, electrical equipment, and steel industries.
Candidate GPTs
I analyze chemicalization, electrification, the internal combustion engine, and interchangeable manufacture as potential drivers of GPT-style transformations in the IR-2. Of these four, electricity is the prototypical GPT. It is “unanimously seen in the literature as a historical example of a GPT.”43 Electricity is one of three technologies, alongside the steam engine and information and communications (ICT) technology, that feature in nearly every article that seeks to identify GPTs throughout history.44 Electrical technologies possessed an enormous scope for improvement, fed into a variety of products and processes, and synergized with many other streams of technological development. Empirical efforts to identify GPTs with patent data provide further evidence of electricity as a GPT in this period.45
Like advances in electricity, clusters of innovations in chemicals and the internal combustion engine not only spurred the rapid growth of new industries but also served as a potential source of GPT trajectories. Historians of technology pick out chemicalization, alongside electrification, as one of two central processes that transformed production routines in the early twentieth century.46 Historical patent data confirm that chemical inventions could influence a wide variety of products and processes.47
In line with GPT classification schemes by other scholars, I also evaluate the internal combustion engine as a candidate GPT, with the potential to replace the steam engine as a prime mover of many industrial processes.48 After its introduction, many believed that the internal combustion engine would transform a range of manufacturing processes with smaller, divisible power units.49
Lastly, I examine the advance of interchangeable manufacture, spurred by innovations in machine tools, as a candidate GPT in this period. Though the machine tool industry was neither new nor especially fast-growing, it did play a central role in extending the mechanization of machine-making first incubated in the IR-1. The diffusion of interchangeable manufacture, or the “American system,” owed much to advances in turret lathes, milling machines, and other machine tools that improved the precision of cutting and shaping metals. Rosenberg’s seminal study of “technological convergence” between the American machine tool industry and metal-using sectors highlighted how innovations in metalworking machines transformed production processes across a wide range of industries.50 Following Rosenberg’s interpretation, historians recognize the nexus of machine tools and mechanization as one of the key technological trajectories during this period.51
Sources of LS and GPT Trajectories
I aimed to include as many candidate technological drivers as possible, provided that the technological developments credibly met the criteria of a leading sector or GPT.52 All candidate leading sectors and GPTs I study in this period were flagged in multiple articles or books that explicitly identified leading sectors or GPTs in the IR-2 period, which helped provide an initial filter for selection. This allows for a good test of the GPT diffusion mechanism against the LS product cycles mechanism.53 This sorting process is an important initial step for evaluating the two mechanisms, though a deeper excavation of the historical evidence is required to determine whether the candidates actually made the cut.
table 4.1 Key Sources of Technological Trajectories in the IR-2
Candidate Leading Sectors
Candidate GPTs
Steel industry
Interchangeable manufacture
Electrical equipment industry
Electrification
Chemicals industry
Chemicalization
Automobile industry
Internal combustion engine
There is substantial overlap between the candidate GPTs and leading sectors in the IR-2, as reflected in table 4.1, but two key distinctions are worth emphasizing. First, one difference between the candidate GPTs and leading sectors is the inclusion of machine tools in the former category. The international relations scholarship on leading sectors overlooks the impact of machine tools in this period, possibly because the industry’s total output did not rank among the largest industries, and also because innovation in machine tools was relatively incremental.54 One survey of technical development in machine tools from 1850 to 1914 described the landscape as “essentially a series of minor adaptations and improvements.”55 Relatedly, the steel industry, commonly regarded as an LS, is not considered a candidate GPT. Under the GPT mechanism, innovations in steel are bound up in a GPT trajectory driven by advances in machine tools.
Second, even though some technological drivers, such as electricity, are considered both candidate leading sectors and candidate GPTs, there are different interpretations of how developments in these technological domains translated into an economic power transition. In the case of new electrical discoveries, control over market share and exports in the electrical equipment industry represents the LS trajectory, whereas the gradual spread of electrification across many industries stands in for the GPT trajectory. Two trajectories diverge in a yellow wood, and the case study evidence will show which one electricity traveled.56
GPT vs. LS Trajectories in the IR-2
Equipped with a better grasp of the possible technological drivers in the IR-2, I follow the same procedures used in the previous chapter to assess the validity of the GPT and LS mechanisms.
OBSERVABLE IMPLICATIONS RELATED TO THE IMPACT TIMEFRAME
GPT diffusion and LS product cycles present two competing interpretations of the IR-2’s impact timeframe. The LS mechanism expects growth associated with radical technological breakthroughs to be explosive in the initial stages. Under this view, off the back of major breakthroughs such as the first practical electric dynamo (1871), the modern internal combustion engine (1876), and the successful synthesis of indigo dye (1880), new leading sectors took off in the 1870s and 1880s.57 Then, according to the expected timeline of the LS mechanism, these new industries stimulated substantial growth in the early stages of their development, bringing about a pre–World War I upheaval in the industrial balance of power.58
The GPT trajectory gives a different timeline for when productivity benefits from major technological breakthroughs were realized on an economy-wide scale. Before stimulating economy-wide growth, the candidate GPTs that emerged in the 1880s—tied to advances in electricity, chemicals, and the internal combustion engine—required many decades of complementary innovations in application sectors and human capital upgrading. These candidate GPTs should have contributed only modestly to the industrial rise of the United States before World War I, with impacts, if any, materializing toward the very end of the period.
Critically, one candidate GPT should have produced substantial economic effects during this period. Unlike other GPT trajectories, interchangeable manufacture had been incubated by earlier advances in machine tools, such as the turret lathe (1845) and the universal milling machine (1861).59 Thus, by the late nineteenth century, interchangeable manufacturing methods should have diffused widely enough to make a significant impact on US industrial productivity.
OBSERVABLE IMPLICATIONS RELATED TO THE PHASE OF RELATIVE ADVANTAGE
When spelling out how the IR-2 produced an economic power transition, the two mechanisms also stress different phases of technological change. According to the LS mechanism, Britain’s industrial prominence waned because it lost its dominance of innovation in the IR-2’s new industries. The United States and Germany benefited from monopoly profits accrued from being lead innovators in electrical equipment, chemical production, automobiles, and steel. In particular, Germany’s industrial rise in this period draws a disproportionate share of attention. Many LS accounts attribute Germany’s rise to its dominance of innovations in the chemical industry, “the first science-based industry.”60 Others emphasize that the American global lead in the share of fundamental innovations after 1850 paved the way for the United States to dominate new industries and become the leading economy in the IR-2.61
The GPT mechanism has different expectations regarding the key determinant of productivity differentials. Where innovations are adopted more effectively has greater significance than where they are first introduced. According to this perspective, Britain lost its industrial preeminence because the United States was more effective at intensively adopting the IR-2’s GPTs.
OBSERVABLE IMPLICATIONS RELATED TO BREADTH OF GROWTH
Finally, regarding the breadth of growth, the third dimension on which the two mechanisms diverge, the LS trajectory expects that a narrow set of modernized industries drove productivity differentials, whereas the GPT trajectory holds that a broad range of industries contributed to productivity differentials. The US growth pattern serves as the best testing ground for these diverging predictions, since the United States overtook Britain as the economic leader in this period.
Table 4.2 Testable Predictions for the IR-2 Case Analysis
Prediction 1: LS (impact timeframe)
The steel, electrical equipment, chemicals, and/or* automobile industries made a significant impact on the rise of the United States to productivity leadership before 1914.
Prediction 1: GPT
Electrification, chemicalization, and/or the internal combustion engine made a significant impact on the rise of the United States to productivity leadership only after 1914.The extension of interchangeable manufacture made a significant impact on the rise of the United States to productivity leadership before 1914.
Prediction 2: LS (phase of relative advantage)
Innovations in the steel, electrical equipment, chemicals, and/or automobile industries were concentrated in the United States.German and American advantages in the production and export of electrical equipment, chemical products, automobiles, and/or steel were crucial to their industrial superiority.
Prediction 2: GPT
Innovations in machine tools, electricity, chemicals, and/or the internal combustion engine were not concentrated in the United States.
American advantages in the diffusion of interchangeable manufacture were crucial to its productivity leadership.
Hypothesis 3: LS (breadth of growth)
Productivity growth in the United States was limited to the steel, electrical, chemicals, and/or automotive industries.
Hypothesis 3: GPT
Productivity growth in the United States was spread across a broad range of industries linked to interchangeable manufacture.
*The operator “and/or” links all the candidate leading sectors and GPTs because it could be the case that only some of these technologies drove the trajectories of the period.
The two explanations hold different views about how technological disruptions produced an economic power transition, related to the impact timeframe of new advances, the phase of technological change that yields relative advantages, and the breadth of technology-fueled growth. Based on the differences between the LS and GPT mechanism across these dimensions, I derive three sets of diverging predictions for how technological changes contributed to relative shifts in economic productivity during this period. Table 4.2 collects these predictions, which structure the case analysis in the following sections.
Impact Timeframe: Gradual Gains vs. Immediate Effects from New Breakthroughs
The opening move in assessing the LS and GPT mechanisms is determining when the IR-2’s eye-catching technological advances actually made their mark on leading economies. Tracking the development timelines for all the candidate leading sectors and GPTs of the IR-2 produces two clear takeaways. First, innovations related to electricity, chemicals, and the internal combustion engine did not make a significant impact on US productivity leadership until after 1914. Second, advances in machine tools and steel—the remaining candidate GPT and leading sector, respectively—contributed substantially to US economic growth before World War I; thus, their impact timeframes fit better with when the United States overtook Britain as the preeminent economic power.
DELAYED TIMELINES: CHEMICALS, ELECTRICITY, AND THE INTERNAL COMBUSTION ENGINE
Developments in chemicals, electricity, and internal combustion provide evidence against the LS interpretation. If the LS mechanism was operational in the IR-2, advances in chemicals should have made a significant impact on US productivity leadership before World War I.62 Yet, in 1914, the United States was home to only seven dye-making firms.63 Major US chemicals firms did not establish industrial research laboratories like those of their German counterparts until the first decade of the twentieth century.64 Terry Reynolds, author of a history of the American Institute of Chemical Engineers, concludes, “Widespread use of chemists in American industrial research laboratories was largely a post–World War I phenomenon.”65 Thus, it is very unlikely that chemical innovations made a meaningful difference to growth differentials between the United States and Britain before 1914.
At first glance, the growth of the German chemical industry aligns with the LS model’s expectations. Germany was the first to incorporate scientific research into chemical production, resulting in the synthesis of many artificial dyes before 1880.66 Overtaking Britain in leadership of the chemical industry, Germany produced 140,000 tons of dyestuffs in 1913, more than 85 percent of the world total.67
While Germany’s rapid growth trajectory in synthetic dyes was impressive, the greater economic impacts of chemical advances materialized after 1914 through a different pathway: “chemicalization,” or the spread of chemical processes across ceramics, food-processing, glass, metallurgy, petroleum refining, and many other industries.68 Prior to key chemical engineering advances in the 1920s, industrial chemists devoted limited attention to unifying principles across the manufacture of different products. The rapid expansion of chemical-based industries in the twentieth century owed more to these later improvements in chemical engineering than earlier progress in synthetic dyes.69 Ultimately, these delayed spillovers from chemicalization were substantial, as evidenced by higher growth rates in the German chemical industry during the interwar period than in the two decades before World War I.70
Electrification’s impact timeframe with respect to US productivity growth mirrored that of chemicalization. Scholarly consensus attributes the US productivity upsurge after 1914 to the delayed impact of the electrification of manufacturing.71 From 1880 to 1930, power production and distribution systems gradually evolved from shaft and belt drive systems driven by a central steam engine or water wheel to electric unit drive, in which electric motors powered individual machines. Unit drive became the predominant method in the 1920s only after vigorous debates in technical associations over its relative merits, the emergence of large utilities that improved access to cheap electricity, and complementary innovations, like machine tools, that were compatible with electric motors.72
Quantitative indicators also verify the long interval between key electrical advances and electrification’s productivity boost. Economic geographer Sergio Petralia has investigated the causal relationship between adoption of electrical and electronic (E&E) technologies, operationalized as E&E patenting activity in individual American counties and the per-capita growth of those counties over time. One of his main findings is that the effects of E&E technology adoption on growth are not significant prior to 1914.73 This timeline is confirmed by a range of other metrics, including the energy efficiency of the American economy, electric motors’ share of horsepower in manufacturing, and estimates of electricity’s total contribution to economic growth.74
The diffusion of internal combustion engines across application sectors was also slow. Despite its initial promise, the internal combustion engine never accounted for more than 5 percent of the generation of total horsepower in US manufacturing from 1869 to 1939.75 In 1900, there were only eight thousand cars in the entire United States, and the U. motor vehicle industry did not overtake its French competitor as the world’s largest until 1904.76 Furthermore, the turning point for the mass production of automobiles, Ford’s installation of a moving assembly line for making Model Ts, did not occur until 1913.77
KEY TIMINGS: MACHINE TOOLS AND STEEL
When assigning credit to certain technologies for major upheavals in global affairs, awe of the new often overwhelms recognition of the old. Based on the previous analysis, it is unlikely that new breakthroughs in electricity, chemicals, and internal combustion fueled the economic power transition that transpired in this period. Instead, careful tracing reveals the persevering impact of earlier developments in machine tools.78 During the IR-2, technical advances in machine tools were incremental, continuous improvements that helped disseminate transformative breakthroughs from the mid-nineteenth century, such as the turret lathe and the universal milling machine.79
Profiles of key application sectors and quantitative indicators validate the GPT mechanism’s expected impact timeframe for machine tools. Marking 1880 as the date when “the proliferation of new machine tools in American industry had begun to reach torrential proportions,” Rosenberg outlines how three application sectors—sewing machines, bicycles, and automobiles—successively adopted improved metal-cutting techniques from 1880 to 1910.80 As the American system took hold, the number of potential machine tool users multiplied 15-fold, from just 95,000 workers in 1850 to almost 1.5 million in 1910.81 Patenting data identify the last third of the nineteenth century as the period when extensive technological convergence characterized the machine tool industry and application sectors.82
FIGURE 4.3. Technological Impact Timeframes in the IR-2. Note: US chemical production, horsepower from electric central stations, and machine intensity over time. Source: Murmann 2003; US Census Bureau 1975.
Figure 4.3 depicts the diverging impact timeframes of interchangeable manufacturing methods, electrification, and chemicalization. Machine intensity substantially increased from 1890 to 1910, as measured by horsepower installed per persons employed in manufacturing. By contrast, the United States did not experience significant increases in electrical and chemical production until after 1910.
Of all the candidate leading sectors, the steel industry best fits the expectations of the LS mechanism regarding when industries transformed by radical innovations stimulated growth in the rising powers. Just as the 1780s were a period when the technological conditions for cotton production were transformed, the mid-nineteenth century featured major breakthroughs in the steel industry that allowed for the mass production of steel, such as the Siemens-Martin open-hearth furnace (1867) and Bessemer converter (1856).83 Over the course of the IR-2 period, the United States and Germany quickly exploited these breakthroughs in steelmaking to massively boost steel production.
The overtaking of Britain by both Germany and the United States in total steel production by the early 1890s matches the timeline of Britain’s overall economic decline.84 Paul Kennedy cites Germany’s booming steel output as a key factor driving its industrial rise; by 1914, German steel output was larger than that of Britain, France, and Russia combined.85 Likewise, US steel output grew from one-fifth of British production in 1871 to almost five times more than British steel output in 1912.86 Given these impressive figures, the next section investigates the American and German advantages in steel production in further detail.
Phase of Relative Advantage: The American System’s Diffusion
The second dimension on which the GPT and LS trajectories differ relates to the phase of technological change that accounted for the relative success of the United States in the IR-2. Cross-country historical evidence on the IR-2’s technological drivers illustrates that the United States had true comparative advantages over other advanced economies that were rooted in its absorption and diffusion capabilities.
INNOVATION CLUSTERING IN STEEL, ELECTRICITY, CHEMICALS, AND/OR MOTOR VEHICLES?
In electricity, industrial powers fiercely contested innovation leadership as the United States, Germany, Great Britain, and France all built their first central power stations, electric trams, and alternating current power systems within a span of nine years.87 However, the United States clearly led in diffusing these systems: US electricity production per capita more than doubled that of Germany, the next closest competitor, in 1912. Along this metric of electrification, Britain’s level was just 20 percent of the US figure.88
To be clear, Britain fell behind in adopting electrification, even though it introduced some of the most significant electrical innovations.89 In 1884, for example, British inventor Charles Parsons demonstrated the first steam turbine for practical use, an essential step for commercializing electric power, but this technology was more rapidly and widely adopted in other countries.90 The British Institution of Electrical Engineers aptly captured this phenomenon in an 1892 resolution: “Notwithstanding that our countrymen have been among the first in inventive genius in electrical science, its development in the United Kingdom is in a backward condition, as compared with other countries, in respect of practical application to the industrial and social requirements of the nation.”91
In chemicals, the achievements of both the US and German chemical industries suggest that no single country monopolized innovation in this sector. Germany’s synthetic dye industry excelled not because it generated the initial breakthroughs in aniline-violet dye processes—in fact, those were first pioneered in Britain—but because it had perfected these processes for profitable exploitation.92 Similar dynamics characterized the US chemical industry.93
In most cases, the United States was not the first to introduce major innovations in leading sectors. Many countries introduced major innovations in chemicals, electricity, motor vehicles, and steel during this period (table 4.3).94 Across the four candidate leading sectors, American firms pioneered less than 30 percent of the innovations. Contrary to the propositions of the LS mechanism, innovations in steel, electricity, chemicals, and motor vehicles were spread across the leading economies.
table 4.3 Geographic Distribution of Major Innovations in Leading Sectors, 1850–1914
Chemicals
Electricity
Motor Vehicles
Steel
France
2
1
1
1
Germany
3
3
3
0
Great Britain
1
3
1
1
United States
2
3
1
0
Various other countries
0
1
0
2
Sole US share
25%
27%
17%
0%
Source: Van Duijn 1983, 176–79 (compilation of 160 innovations introduced during the nineteenth and twentieth centuries).
Moreover, the limited role of electrical and chemical exports in spurring American growth casts further doubt on the significance of monopoly profits from being the first to introduce new advances.95 The British share of global chemical exports almost doubled the US share in 1913.96 Overall, the United States derived only 8 percent of its national income from foreign trade in 1913, whereas the corresponding proportion for Britain was 26 percent.97 Even though the United States was the quickest to electrify its economy, Germany captured around half of the world’s exports in electrical products.98
If monopoly profits from innovation clustering in any leading sector propelled the industrial rise of the United States and Germany, it would be the steel industry. Both nations made remarkable gains in total steel output over this period, and scholars commonly employ crude steel production as a key indicator of British decline and the shifting balance of industrial power in the decades before World War I.99 Thus, having established the delayed impact of the electrical, chemical, and automobile industries in this period, the steel industry takes on an especially large burden for the LS mechanism’s explanatory power in this period.
Yet Britain capitalized on many major innovations in steelmaking, including the Talbot furnace, which became essential to producing open-hearth steel.100 Moreover, trade patterns reveal that Britain still held a comparative advantage in the export of steel between 1899 and 1913.101 How to square this with Germany’s dominance in total steel output?
The prevailing wisdom takes total steel output figures to stand for superior American and German technological know-how and productivity.102 In truth, new steelmaking processes created two separate steel industries. Britain shifted toward producing open-hearth steel, which was higher in quality and price. According to the British Iron Trade Association, Britain produced about four times more open-hearth steel than Germany in 1890.103 On the other hand, Germany produced cheap Thomas steel and exported a large amount at dumping prices. In fact, some of Germany’s steel exports went to Britain, where they were processed into higher-quality steel and re-exported.104 In sum, this evidence questions what one scholar deems “the myth of the technological superiority and outstanding productivity of the German steel industry before and after the First World War.”105
AMERICAN MACHINE TOOLS—GPT DIFFUSION ADVANTAGE
Though new industries like electricity and chemicals hog much of the spotlight, developments in machine tools underpin the most important channel between differential rates of technology adoption and the IR-2’s economic power transition. After noting the importance of the electrical and chemical industries during the period, British historian Eric Hobsbawm elevates the importance of machine tools: “Yet nowhere did foreign countries—and again chiefly the USA—leap ahead more decisively than in this field.”106
In line with the expectations of GPT diffusion theory, comparative estimates confirm a substantial US lead in mechanization in the early twentieth century. In 1907, machine intensity in the United States was more than two times higher than rates in Britain and Germany.107 In 1930, the earliest year for which data on installed machine tools per employee are available, Germany lagged behind the United States in installed machine tools per employee across manufacturing industries by 10 percent, with a significantly wider gap in the tools most crucial for mass production.108
This disparity in mechanization was not rooted in the exclusive access of the United States to special innovations in machine tools. In terms of quality, British machine tools were superior to their American counterparts throughout the IR-2 period.109 German firms also had advantages in certain fields like sophisticated power technology.110 Rather, the distinguishing feature of the US machine tool industry was excellence in diffusing innovations across industries.111 Reports by British and German study trips to the United States provide some of the most detailed, reliable accounts of transatlantic differences in manufacturing methods. German observers traveled to the United States to learn from their American competitors and eventually imitate American interchangeable manufacturing methods.112 British inspection teams reported that the US competitive edge came from the “adaptation of special apparatus to a single operation in almost all branches of industry”113 and “the eagerness with which they call in the aid of machinery in almost every department of industry.”114
Fittingly, one of the most colorful denunciations of American innovation capacity simultaneously underscored its strong diffusion capacity. In an 1883 address to the American Association for the Advancement of Science, Henry Rowland, the association’s vice president, denigrated the state of American science for its skew toward the commercialization of new advances. Rowland expressed his disgust with media representations that upheld the “obscure American who steals the ideas of some great mind of the past, and enriches himself by the application of the same to domestic uses” over “the great originator of the idea, who might have worked out hundreds of such applications, had his mind possessed the necessary element of vulgarity.”115 Yet, it was America’s diffusion capacity—in all its obscurity and vulgarity—that sustained its growth to economic preeminence.
Breadth of Growth: The Wide Reach of Interchangeable Manufacture
What were the sources of American productivity growth in the IR-2? The pattern of American economic growth is most pertinent to investigate because the United States overtook Britain in productivity leadership during the IR-2. Regarding the breadth of economic growth, the LS trajectory expects that American productivity growth was concentrated in a narrow set of modernized industries, whereas the GPT trajectory holds that American productivity growth was dispersed across a broad range of industries. Sector-level estimates of total factor productivity (TFP) growth provide useful evidence to assess these diverging propositions.
WIDESPREAD PRODUCTIVITY GROWTH
The historical data support GPT diffusion theory’s expectation of pervasive US productivity growth. John Kendrick’s detailed study of US productivity growth in this period depicts a relatively balanced distribution. Among the industries studied, nearly 60 percent averaged between 1 and 3 percent increases in output per labor-hour from 1899 to 1909.116 Broad swathes of the US economy, outside of the leading sectors, experienced technological change. For instance, the service sector, which included segments of the construction, transport, wholesale, and retail trade industries, played a key role in the US capacity to narrow the gap with Britain in productivity performance.117
R&D-centric sectors were not the primary engines of US growth. In a recent update to Kendrick’s estimates, a group of researchers estimated how much of US productivity growth was driven by “great inventions sectors,” a designation that roughly corresponds to this chapter’s candidate leading sectors.118 They found that these sectors accounted for only 29 percent of U.S. TFP growth from 1899–1909.119 Despite employing 40 percent of all research scientists in 1920, the chemical industry was responsible for only 7 percent of US TFP growth throughout the following decade.120
MACHINE TOOLS AND BROADLY DISTRIBUTED PRODUCTIVITY GROWTH
Broad-based productivity growth in the US economy does not necessarily mean that a GPT was at work. Macroeconomic factors or the accumulation of various, unconnected sources of TFP growth could produce this outcome. Therefore, if the GPT trajectory captures the breadth of growth in the IR-2, then the historical evidence should connect broadly distributed productivity growth in the United States to developments in machine tools.
The extension of the American system boosted productivity in a wide range of sectors. Applications of this system of special tools reshaped the processes of making firearms, furniture, sewing machines, bicycles, automobiles, cigarettes, clocks, boots and shoes, scientific instruments, typewriters, agricultural implements, locomotives, and naval ordnance.121 Its influence covered “almost every branch of industry where articles have to be repeated.”122 Per a 1930 inventory of American machine tools, the earliest complete survey, nearly 1.4 million metalworking machines were used across twenty industrial sectors.123 In his seminal study of American productivity growth during this period, Kendrick identifies progress in “certain types of new products developed by the machinery and other producer industries [that] have broad applications across industry lines” as a key source of the “broad, pervasive forces that promote efficiency throughout the economy.”124
The breadth of productivity spillovers from machine tools was not boundless. Machine-using industries constituted a minority of the manufacturing industries, which themselves accounted for less than one-quarter of national income.125 However, users of new machine tools extended beyond just manufacturing industries. Technologically intensive services, such as railroads and steam transportation, also benefited significantly from improved metalworking techniques.126 In agriculture, specialized machine tools helped advance the introduction of farm machinery such as the reaper, which revolutionized agricultural productivity.127
In describing how machine tools served as a transmission center in the US economy, Rosenberg describes the industry as a pool of skills and technical knowledge that replenishes the economy’s machine-using sectors—that is, an innovation that addresses one industry’s problem gets added to the pool and becomes available, with a few modifications, for all technologically related industries.128 As sales records from leading machine tool firms show, many application sectors purchased the same type of machine. In 1867, Brown and Sharpe Manufacturing Company sold the universal milling machine, just five years after its invention, not only to machinery firms that made tools for a diverse range of industries but also to twenty-seven other firms that produced everything from ammunition to jewelry.129 In this way, the machine tool industry functioned, in Rosenberg’s words, as “a center for the acquisition and diffusion of new skills and techniques in a machinofacture type of economy.”130
Indeed, advances in machine tools had economy-wide effects. The social savings method estimates how much a new technology contributed to economic growth, compared to a counterfactual situation in which the technology had not been invented.131 Referencing this method to differentiate between the impacts of new technologies in this period, economic historian Joel Mokyr puts forward the American system of manufacturing as the most important:
From a purely economic point of view, it could be argued that the most important invention was not another chemical dye, a better engine, or even electricity.… There is one innovation, however, for which “social savings” calculations from the vantage point of the twentieth century are certain to yield large gains. The so-called American System of manufacturing assembled complex products from mass-produced individual components. Modern manufacturing would be unthinkable without interchangeable parts.132
Institutional Complementarities: GPT Skill Infrastructure in the IR-2
With confirmation that the pattern of technological change in the IR-2 is better characterized by the GPT trajectory, the natural next step is to probe variation among leading economies in adapting to this trajectory. Why was the United States more successful than Britain and Germany in adapting to the demands of interchangeable manufacture? According to GPT diffusion theory, the historical evidence should reveal that the US edge was based on education and training systems that broadened and systematized mechanical engineering skills. These institutional adaptations would have resolved two key bottlenecks in the spread of interchangeable manufacture: a shortage of mechanical engineering talent and ineffective coordination between machine tool producers and users.
Widening the Base of Mechanical Engineers
Which institutions for skill formation were most central to the ability of the United States to take advantage of new advances in machine tools? Established accounts of economic rivalry among great powers in the IR-2 focus on skills linked to major innovations in new, science-based industries. Emphasizing Germany’s advantage in training scientific researchers, these studies attribute Germany’s technological success in this period to its investments in R&D facilities and advanced scientific and technical education.133 Such conclusions echo early-twentieth-century British accounts of Germany’s growing commercial prowess, which lauded German higher education for awarding doctorates in engineering and its qualitative superiority in scientific research.134
American leadership in the adoption of interchangeable manufacturing methods was beholden to a different set of institutions for skill formation. Progress in this domain did not depend on new scientific frontiers and industrial research laboratories.135 In fact, the United States trailed both Britain and Germany in scientific achievements and talent.136 Widespread mechanization in the United States rested instead on a broad base of mechanical engineering skills.
Alongside the development of more automatic and precise machine tools throughout the nineteenth century, this new trajectory of mechanization demanded more of machinists and mechanical engineers. Before 1870, US firms relied on informal apprenticeships at small workshops for training people who would design and use machine tools.137 At the same time, engineering education at independent technical schools and traditional colleges and universities did not prioritize mechanical engineers but were mostly oriented toward civil engineering.138 Yet craft-era methods and skills were no longer sufficient to handle advances that enhanced the sophistication of machine tools.139 Thus, in the mid-eighteenth century, the US potential for mechanization was significantly constrained by the need for more formal technical instruction in mechanical engineering.
Over the next few decades, advances on three main fronts met this need for a wider pool of mechanical engineering expertise: land-grant schools, technical institutes, and standardization efforts. In 1862, the US Congress passed the first Morrill Land-Grant Act, which financed the creation of land-grant colleges dedicated to the agricultural and mechanical arts. Although some of these schools offered low-quality instruction and initially restricted their mission to agricultural concerns, the land-grant funds also supported many important engineering schools, such as the Massachusetts Institute of Technology (MIT) and Cornell University.140 The number of US engineering schools multiplied from six in 1862, when the Morrill Act was passed, to 126 in 1917.141 These schools were especially significant in widening the base of professional mechanical engineers. In 1900, out of all students pursuing mechanical engineering at US higher education institutions, 88 percent were enrolled in land-grant colleges.142
The establishment of technical institutes also served demands for mechanical engineering training. Pure technical schools like the Worcester Polytechnic Institute, founded in 1868, and the Stevens Institute of Technology, founded in 1870, developed mechanical engineering curricula that would become templates for engineering programs at universities and colleges.143 Embedded with local and regional businesses, technical institutes developed laboratory exercises that familiarized students with real-world techniques and equipment. In this respect, these institutes and land-grant colleges “shared a common belief in the need to deliver a practice-oriented technical education.”144
Another significant development in the spread of mechanical engineering knowledge was the emergence of professional engineering societies that created industrial standards. The most prominent of these were the American Society of Mechanical Engineers (ASME), founded in 1880, the American Section of the International Association for Testing Materials, set up in 1898, and the Franklin Institute, which became America’s leading technical society around the start of the IR-2.145 As these associations coordinated to share best practices in mechanical engineering, they improved knowledge flows between the machine tool industry and application sectors.146 Standardization in various machine processes and components, such as screw threads, helped spread mechanization across disparate markets and communities.147
It should be emphasized that these efforts were effective in producing the skills and knowledge necessary for advancing mechanization because they broadened the field of mechanical engineering. Mechanical engineering instruction at land-grant schools and technical institutes and through professional associations allowed for more students to become “average engineers,” as opposed to “the perpetuation of a self-recognized elite.”148 Recent research finds that this diffused engineering capacity produced enduring benefits for American industrialization. By collecting granular data on engineering density for the United States at the county level, William Maloney and Felipe Caicedo capture the engineering talent spread across various US counties in 1880 and parse the effect of engineering capacity on industrial outcomes decades later. They find that there is a statistically significant, positive relationship between the level of engineering density in 1880 and the level of industrialization decades later.149
The Comparative Advantage of the United States over Britain and Germany in GPT Skill Infrastructure
Both Britain and Germany fell short of the US standard in GPT skill infrastructure. For Britain, the key gap was in the supply of mechanical engineering talent. British educational institutions and professional bodies fiercely guarded the apprenticeship tradition for training mechanical engineers.150 For instance, the University of Oxford did not establish an engineering professorship until 1908.151 Meanwhile, American engineers systematically experimented with machine redesigns, benefiting from their training at universities and technical institutes.
These diverging approaches resulted in stark differences in skill formation. In 1901, probably around 2,600 students were enrolled in full-time higher technical education in the United Kingdom.152 Limiting this population to those in their third or fourth year of full-time study—an important condition because many UK programs, unlike German and American institutions, did not progress beyond two years of study—leaves only about 400 students.153 By comparison, in mechanical engineering programs alone, the United States in 1900 had 4,459 students enrolled in higher technical education.154 Controlling for population differences, the United States substantially outpaced Britain in engineering density, as measured by the number of university-educated engineers per 100,000 male laborers.155
Germany developed a more practical and accessible form of higher technical education than Britain. From 1870 to 1900, enrollments in the German technische Hochschulen increased nearly fourfold, from 13,674 to 32,834 students.156 Alongside the technische Mittelschulen (technical intermediate schools comparable to American industrial trade schools and lower-level engineering colleges), the technische Hochschulen cultivated a broad base of mechanical engineers.157 Germany’s system of technical education attracted admirers from around the world. Some went there to study in the schools, and others went to study how the school system worked, with the aim of borrowing elements of the German model.158
Germany’s problems were with weak linkages between mechanical engineering education and industrial applications. Key German standards bodies and technical colleges prioritized scientific and theoretical education at the expense of practical skills—a trend “most pronounced in mechanical engineering.”159 According to an expert on German standard-setting in this period, “no national standards movement was inaugurated in [the machine industry] until after the outbreak of [World War I].”160 German experts on engineering education, intent on reforming technical instruction to get engineers more experience with factor organization and project management in the field, recommended, for example, that practical training courses be offered in partnerships with engineering associations.161 Articles in the Zeitschrift des Vereines Deutscher Ingenieure (Journal of the Association of German Engineers) lamented that the technische Hochschulen and technical universities were not equipping students with practical skills to operate in and manage factories and workshops.162 These issues slowed Germany’s incorporation of interchangeable parts and advanced machine tools.
A report by Professor Alois Riedler of the Technical University of Berlin, who was commissioned by the Prussian Ministry of Education to tour American engineering schools in the 1890s, illustrates the differences in engineering education between the United States and Germany. According to Riedler, extensive practical training and experience with shop and laboratory applications were distinctive features of an American engineering education. To substantiate differences in practical instruction between engineering departments in the two countries, Riedler analyzed the time allocated to theoretical and practical instruction across four-year courses of study.163 Compared to their German peers, American students spent far more time on exercises in mechanical technical laboratories and other types of practical training (figure 4.4). In the Technische Universität Berlin (Technical University Berlin), practical exercises in the laboratory and shop accounted for less than 6 percent of total instruction time over a four-year course of study.164 In contrast, engineering students at Cornell University spent more than one-third of their course engaged in laboratory study and shopwork. As a consequence of reports by Riedler and others, German institutions began establishing laboratories for mechanical engineering around 1900.165
FIGURE 4.4. Comparison of Curricula at German and American Engineering Schools (1893). Source: US Bureau of Education (BOE) 1895, 684–86. Note: In this BOE report, the German schools are labeled Technological University in Austria, Technological University in Prussia, and Technological University in South Germany. A reasonable assumption, informed by background research, is that these refer to Technical University Wien, Technical University Berlin, and Technical University of Munich, respectively. Though TU Wien is in Austria, it is used to illustrate trends in German engineering education because many German schools saw it as an influential model.
It should be made clear that, in the United States, institutional adaptations to new opportunities presented by interchangeable manufacture were not rooted in cultivating highly skilled scientific talent. The best and brightest American scientists furthered their education at European universities.166 Even proponents of American engineering education concluded that “strictly scientific and intellectual education in American technological schools” did not even match “the average of a secondary industrial school” in Germany.167 According to one study conducted by the National Association of German-American Technologists, an organization that regularly circulated ideas between the two countries, German technical institutes held an edge over their US peers in research on different technologies in mechanical engineering.168
The deeper roots of US institutions’ greater effectiveness at adapting to the skill formation needs of interchangeable manufacture cannot be fully explored here. The legacy of the Morrill Act certainly looms large, as do the contributions of a diverse set of institutional adaptations unrelated to that groundbreaking federal policy, including independent centers like the Franklin Institute, technical high schools, professional associations, and specialized engineering programs initiated at preexisting universities.169 Other potential sources for the US advantage in GPT skill infrastructure include its openness to foreign technicians and the unique challenges and culture of the American frontier.170
LS-Based Theories and Chemical Engineering
Analyzing the education and training systems for chemical advances provides a secondary test to determine which institutions are most apt to bring national success in technological revolutions.171 LS accounts typically point to Germany’s innovation capacity as the key determinant of its competitiveness in chemicals, especially in the key segment of synthetic dye production.172 To extend this lead in synthetic dyes, Germany profited from leading industrial research labs and scientific education institutions, which employed the world’s top academic chemists and produced about two-thirds of the world’s chemical research.173
By comparison, the US capacity to innovate in chemicals was weak. From 1901 to 1930, only one American researcher received a Nobel Prize in Chemistry, while German and British researchers captured almost three-fourths of the Nobel Prizes in Chemistry in that span.174 In 1899, German publications accounted for half of all citations in American chemical journals, essentially double the share credited to American publications.175 At the same time, American scholarship barely registered in Europe-based chemistry journals, where the best research was published. According to one analysis of references in Annual Reports on the Progress of Chemistry, an authoritative British review journal, American publications accounted for only 7 percent of the citations in 1904.176
As was the case with machine tools, effective adaptation to new chemical technologies in the United States rested on a different set of institutional competencies. Despite trailing Germany in chemical breakthroughs and top chemists, the United States pioneered a chemical engineering discipline that facilitated the gradual chemicalization of many industries. A crucial step in this process was the emergence of unit operations, which broke down chemical processes into a sequence of basic operations (for example, condensing, crystallizing, and electrolyzing) that were useful to many industries, including ceramics, food processing, glass, metallurgy, and petroleum refining.177 American institutions of higher education, most notably MIT, quickly adopted the unit operations model and helped cultivate a common language and professional community of chemical engineering.178 As Rosenberg and Steinmueller conclude, “American leadership in introducing a new engineering discipline into the university curriculum, even at a time when the country was far from the frontier of scientific research, was nowhere more conspicuous than in the discipline of chemical engineering early in the 20th century.”179
In contrast, Germany was slow to develop the infrastructure for supporting chemical engineers. Up through the interwar period, the chemical engineering profession “failed to coalesce in Germany.”180 Chemical engineering did not become a distinct academic subject area in Germany until after the Second World War.181 Because German universities did not equip chemists with engineering skills, the burden of training chemists was shifted to firms.182 Additionally, the German chemical industry maintained a strict division of labor between chemists and mechanical engineers. The lack of skill systematization resulted in more secrecy, less interfirm communication, and a failure to exploit externalities from common chemical processes.183
The United States reaped the spoils of technological convergence in chemicalization not just because it trained large numbers of chemical engineers but also because it strengthened university-industry linkages and standardized techniques in chemical engineering.184 Without the connective tissue that promotes information flows between the chemical sector and application sectors, a large base of chemical engineers was insufficient. Britain, for instance, was relatively successful at training chemical engineers during the interwar period; however, the weak links between British educational institutions and industrial actors limited the dissemination of technical knowledge, and the concept of unit operations did not take hold in Britain to the degree that it did in America.185 Additionally, professional engineering associations in the United States, including the American Institute of Chemical Engineers, advanced standardization in the chemical industry—an initiative not imitated in Britain until a decade later.186 Unlike their American peers, it was not until after World War II that British chemical engineers saw themselves as “members of a professional group that shared a broad commonality cutting across the boundary lines of a large number of industries.”187
Since substantial, economy-wide benefits from these chemical breakthroughs did not materialize until after the end of the IR-2 period, it is important to not overstate these points. Nonetheless, tracing which country best exploited chemical innovations through the interwar period can supplement the analysis of institutional complementarities for machine tools.188 Evidence from the coevolution of chemical technologies and skill formation institutions further illustrates how institutional adaptations suited to GPT trajectories differed from those suited to LS trajectories.
Alternative Factors
Like its predecessor, the IR-2 has been the subject of countless studies. Scholars have thoroughly investigated the decline of Britain and the rise of the United States and Germany, offering explanations ranging from immigration patterns and cultural and generational factors to natural resource endowments and labor relations.189 My aim is not to sort through all possible causes of British decline. Rather, I am tracing the mechanisms behind an established connection between the IR-2’s technological breakthroughs and an economic power transition. Thus, the contextual factors most likely to confound the GPT diffusion explanation are those that provide an alternative explanation of how significant technological changes translated into the United States supplanting Britain in economic leadership. Aside from the LS mechanism, which has been examined in detail, the influence of international security threats and varieties of capitalism deserve further examination.
Threat-Based Explanations
How did external threats influence technological leadership in the IR-2? Scholars have argued that US military investment, mobilized against the threat of a major war, was crucial to the development of many GPTs.190 US national armories’ subsidization of the production of small arms with interchangeable parts in the early nineteenth century was crucial, some studies argue, to the diffusion of the American system to other industries in the second half of the century.191
Though firearms production provided an important experimental ground for mechanized production, military support was not necessary to the development of the American system of manufacturing. Questioning the necessity of government funding and subsidies for the spread of the American system, one study credits the development of interchangeable manufacture to four civilian industries: clock manufacturing, ax manufacturing, typewriter manufacturing, and watch manufacturing.192 In particular, the clock industry played a crucial role in diffusing mechanized production practices. More attuned to the dynamics of the civilian economy than the small arms manufacturers, clockmakers demonstrated that interchangeable manufacture could drastically increase sales and cut costs.193 In his definitive study of the history of American interchangeable parts manufacture, David Hounshell concludes that “the sewing machine and other industries of the second half of the 19th century that borrowed small arms production techniques owed more to the clock industry than to firearms.”194
Military investment and government contracting did not provide long-term sources of demand for interchangeable manufacturing methods.195 Over the course of the IR-2, the small arms industry’s contribution to American manufacturing declined, totaling less than 0.3 percent of value added in American industry from 1850 to 1940.196 Thus, arguments centered on military investment neglect that the spread of the American system, not its initial incubation, is the focal point for understanding how the IR-2 catalyzed an economic power transition.197
Another threat-based argument posits that countries that face more external threats than internal rivalries will achieve more technological success.198 In the IR-2 case, however, the United States was relatively isolated from external conflicts, while the United Kingdom and Germany faced many more threats (including each other).199 Moreover, the United States was threatened more by internal rivalries than by external enemies at the beginning of the IR-2, as it had just experienced a civil war.200 This argument therefore provides limited leverage in the IR-2 case.
VoC Explanations
What about the connection between America’s particular type of capitalism and its economic rise? Rooted in the varieties of capitalism (VoC) tradition, one alternative explanation posits that the United States was especially suited to embrace the radical innovations of the IR-2 because it exhibited the characteristics of a liberal market economy (LME). GPT diffusion theory and VoC-based explanations clash most directly on points about skill formation. The latter’s expectation that LMEs like the United States should excel at cultivating general skills could account for US leadership in GPT diffusion during this period.201
The empirical evidence casts doubt on this explanation. In the early twentieth century, the leading nations had fairly similar levels of enrollment rates in elementary and post-elementary education. In 1910, enrollment rates for children ages five to nineteen in the United States was about 12 percent lower than Britain’s rate and only 3 percent higher than Germany’s.202 Years of education per worker increased by essentially the same proportion in both Britain (by a factor of 2.2) and the United States (by a factor of 2.3) between 1870 and 1929.203 In terms of higher education expenditures per capita, the two countries were essentially tied.204 Differences in the formation of general skills cannot account for the technological leadership of the United States in this period.205
Moreover, the degree to which the United States fully embraced the characteristics of LMEs is disputed. In the view of studies that position the United States as a model for managerial capitalism in this period, US industrial governance structures enabled the rise of giant managerialist firms.206 This approach primarily sees America’s rise to industrial preeminence through the most visible actors in the American system of political economy: oligopolies in the automobile, steel, and electrical industries. There was significant diversity, however, in firm structure. Though many giant corporations did grow to take advantage of economies of scale and capital requirements in some mass-produced goods (such as automobiles), networks of medium-sized firms still dominated important segments of these new industries, such as the production of electric motors. One-third of the fifty largest manufacturing plants in the United States made custom and specialty goods.207 From 1899 to 1909, sectors that relied on batch and custom production, including machine tools, accounted for one-third of value added in manufacturing.208 No specific brand of capitalism fulfilled the demands of production across all domains.209
Case-Specific Factors
Another traditional explanation highlights American natural resource abundance as a key factor in transatlantic differences in mechanization. Compared to its European competitors, the benefits to the United States derived from its endowment of natural resources, such as plentiful supplies of timber, biased its manufacturing processes toward standardized production.210 “The American turn in the direction of mass production was natural,” claims one influential study of diverging approaches to mechanization in this period.211
The extent to which natural resource endowments determined transatlantic differences in technological trajectories is disputed. Undeterred by natural resource differences, German engineers and industrialists frequently used American machine tools and imitated US production technology.212 In fact, around the early twentieth century, the level of machine intensity in German industries was catching up to the rate in American industries.213 The US-Germany gap in mechanization was more about Germany’s struggles in proficiently using advanced tools, not the choice of methods shaped by natural resource endowments. Crucially, skill formation and embedded knowledge about working with new machinery influenced the efficient utilization of American capital-intensive techniques.214
Summary
The standard version of the Second Industrial Revolution’s geopolitical aftershocks highlights Germany’s challenge to British power. Germany’s relative economic rise, according to this account, derived from its advantage in industrial research and scientific infrastructure, which enabled it to capture the gains from new industries such as electricity and chemicals. However, a range of indicators emphasize that it was the United States, not Germany, that surpassed Britain in productivity leadership during this period. The US industrial ascent in the IR-2 illustrates that dominating innovation in leading sectors is not the crucial mechanism in explaining the rise and fall of great powers. Britain’s decline was not a failure of innovation but of diffusion. As the renowned economist Sir William Arthur Lewis once mused, “Britain would have done well enough if she merely imitated German and American innovations.”215
Indeed, the IR-2 case further supports the conclusion that capacity to widely diffuse GPTs is the key driver of long-term growth differentials. The US success in broadening its talent base in mechanical engineering proved critical for its relative advantage in adapting machine tools across a broad range of industries. Like all GPT trajectories, this process was a protracted one, but it aligns better with when the United States surpassed Britain in productive leadership than the more dramatic breakthroughs in chemicals, electricity, and automobiles. To further investigate how the LS mechanism breaks down and why the GPT mechanism holds up, we turn to the high-tech competition in the twentieth century between the United States and Japan—or what some label the Third Industrial Revolution.
5 Japan’s Challenge in the Third Industrial Revolution
IN THE TWO previous cases, an industrial revolution preceded a shift in global leadership. Britain established its economic dominance in the early nineteenth century, and the United States took the mantle in the late nineteenth century. During the last third of the twentieth century (1960–2000), the technological environment underwent a transformation akin to the First and Second Industrial Revolutions. A cluster of information technologies, connected to fundamental breakthroughs in computers and semiconductors, disrupted the foundations of many industries. The terms “Third Industrial Revolution” (IR-3) and “Information Age” came to refer to an epochal shift from industrial systems to information-based and computerized systems.1 Amid this upheaval, many thought Japan would follow in the footsteps of Britain and the United States to become the “Number One” technological power.2
Of the countries racing to take advantage of the IR-3, Japan’s remarkable advances in electronics and information technology garnered a disproportionate share of the spotlight. “The more advanced economies, with Japan taking the lead in one industry after another, [were] restructuring their economies around the computer and other high tech industries of the third industrial revolution,” Gilpin wrote.3 In the late 1980s and early 1990s, a torrent of works bemoaned the loss of US technological leadership to Japan.4 In a best-selling book on US-Japan relations, Clyde Prestowitz, a former US trade negotiator, declared, “Japan has … become the undisputed world economic champion.”5
Japan’s dominance in the IR-3’s leading sectors was perceived as a threat to international security and to US overall leadership of the international system.6 Former secretary of state Henry Kissinger and other prominent thinkers warned that Japan would convert its economic strength into threatening military power.7 Per a 1990 New York Times poll, 58 percent of Americans believed that Japan’s economic power was more of a threat to American security than the Soviet Union’s military power.8
Historical precedents loomed over these worries. US policymakers feared that falling behind Japan in key technologies would, like relative declines experienced by previous leading powers, culminate in an economic power transition. Paul Kennedy and other historically minded thinkers likened the US position in the 1980s to Britain’s backwardness a century earlier: two industrial hegemons on the brink of losing their supremacy.9 Often alluding to the LS mechanism, these comparisons highlighted Japan’s lead in specific industries that were experiencing significant technological disruption, such as consumer electronics and semiconductors. As David Mowery and Nathan Rosenberg wrote in 1991, “Rapidly growing German domination of dyestuffs helped to propel that country into the position of the strongest continental industrial power. The parallels to the Japanese strategy in electronics in recent decades are striking.”10
Many voices called for the United States to mimic Japan’s keiretsu system of industrial organization and proactive industrial policy, which they viewed as crucial to the rising power’s success with leading sectors.11 Kennedy’s The Rise and Fall of the Great Powers attributed Japan’s surge in global market shares of high-tech industries to R&D investments and the organizing role of the Ministry of International Trade and Industry (MITI).12 These claims about the basis of Japan’s leadership in the information revolution relied on LS product cycles as the filter for the most important institutional factors.
The feared economic power transition, however, never occurred. To be sure, Japanese firms did take dominant positions in key segments of high-growth industries like semiconductors and consumer electronics. Additionally, the Japanese economy did grow at a remarkable pace, averaging an annual 2.4 percent increase in total factor productivity (TFP) between 1983 and 1991. However, Japan’s TFP growth stalled at an average of 0.2 percent per year in the 1990s—a period known as its “lost decade.” By 2002, the per capita GDP gap between Japan and the United States was larger than it had been in 1980.13 Becoming the world’s leading producer in high-tech industries did not catalyze Japan’s overtaking of the United States as the leading economy.
The IR-3 case is particularly damaging for LS-based explanations. Japan took advantage of the IR-3’s opportunities by cornering the market in new, technologically progressive industries, fulfilling the conditions posited by the LS mechanism for Japan to become the foremost economic power. Yet, as the case study evidence will reveal, an economic power transition did not occur, even though all these conditions were present. The Japanese challenge to American technological leadership in the last third of the twentieth century therefore primarily functions as a deviant, or falsifying, case for the LS mechanism.14
By contrast, the IR-3 case does not undermine the GPT mechanism. Since Japan did not lead the United States in the diffusion of general-purpose information technologies, the conditions for an economic power transition under the GPT mechanism were absent in the IR-3. Since there could be many reasons why an economic power transition does not occur, the absence of a mechanism in a negative case provides limited leverage for explaining how technology-driven economic power transitions occur. Still, the IR-3 case evidence will show that LS theory expects an outcome that does not occur—a US-Japan economic power transition—in part because it fails to account for the relative success of the United States in GPT diffusion. This advantage stemmed from its superior ability to cultivate the computer engineering talent necessary to advance computerization. In that regard, this deviant case can help form better mechanism-based explanations.15
Surprisingly, few scholars have revisited claims that Japan’s leadership in leading sectors meant that it was on its way to economic preeminence.16 Decades of hindsight bring not just perspective but also more sources to pore over. Revised estimates and the greater availability of data help paint a more granular picture of how the US-Japan productivity gap evolved in this period. To narrow down the crucial technological trajectories, I pieced together histories of semiconductors and other key technologies, comparative histories of technological development in the United States and Japan, and general economic histories of the IR-3. In addition, I leveraged bibliometric techniques to estimate the number of universities in both countries that could supply a baseline quality of software engineering education. Surveys on computer utilization by Japanese agencies, presentations on computer science education by Japanese and American analysts at international meetings, documents from the Edward A. Feigenbaum Papers collection, and back issues of Nikkei Computer (日経コンピュータ) at the Stanford University East Asia Library all helped flesh out the state of GPT skill infrastructure in the IR-3.
The evaluation of the GPT and LS mechanisms against historical evidence from the IR-3 proceeds as follows. The chapter first makes clear that a US-Japan economic power transition did not take place. Subsequently, it reviews and organizes the technological breakthroughs of the IR-3 into candidate leading sectors and GPTs. It then examines whether all the components of the GPT or LS mechanism were present. Since the outcome did not occur in this case, it is important to trace where the mechanisms break down. All the aspects of the LS mechanism were present in the IR-3, but the GPT mechanism was not operational because Japan fell behind the United States in diffusing information technologies across a broad range of sectors. Based on this evidence, the next section explains why institutional explanations rooted in LS trajectories are unconvincing. Before turning to alternative factors and explanations, the chapter analyzes whether GPT skill infrastructure was a factor in sustained US technological leadership.
A Power Transition Unfulfilled: Japan’s Rise Stagnates
In a 1983 article for Parade, the Pulitzer Prize–winning journalist David Halberstam described Japan’s industrial ascent as America’s “most difficult challenge for the rest of the century” and “a more intense competition than the previous political-military competition with the Soviet Union.”17 By the end of the century, however, the possibility of Japan displacing the United States as the technological hegemon was barely considered, let alone feared.18 The economic power transition that accompanied the IR-1 and IR-2 did not materialize in this case. Indeed, most indicators presented a clear trend: Japan’s economy catches up in the 1980s, stagnates in the 1990s, and ultimately fails to overtake the US economy in productivity leadership.
GDP PER-CAPITA INDICATORS
During the three decades after 1960, Japan’s economy experienced remarkable growth, reaching a GDP per capita in 1990 that was 81 percent of the US mark that year. In the following ten years, known as Japan’s “lost decade,” Japan’s growth in GDP per capita stalled. By 2007, Japan’s GDP per capita had dropped back down to 73 percent of that of the United States (figure 5.1).
INDUSTRIALIZATION INDICATORS
Comparative industrialization statistics tell a similar story. In terms of global manufacturing output, Japan gained on the United States through the 1970s and 1980s and nearly matched the United States, at 20 percent of global manufacturing output, in the early 1990s. Japan’s share of global manufacturing output subsequently declined to around 10 percent in 2010, while the US share increased in the 1990s and held at 20 percent until 2010.19 In manufacturing industries, Japan’s labor productivity growth from 1995 to 2004 averaged only 3.3 percent, whereas the United States averaged 6.1 percent in the same metric.20
FIGURE 5.1. Japan’s Catch-up to the United States in GDP per Capita Stalls in the 1990s. Note: Real GDP per capita in 2011$ prices. Source: Maddison Project Database, version 2020 (Bolt and van Zanden 2020).
PRODUCTIVITY INDICATORS
Productivity statistics also reveal a general trend of convergence without overtaking. From a total factor productivity of just half that of the United States in 1955, Japan’s TFP grew steadily. By 1991, the productivity gap between the United States and Japan was only 5 percent. As was the case with the GDP per capita and industrialization figures, Japan’s productivity growth then slowed, and the gap between the United States and Japan widened during the 1990s (figure 5.2). Throughout this decade, Japan averaged just 0.2 percent annual TFP growth.21 By 2009, Japan’s TFP dropped to only 83 percent of the US figure. The US-Japan labor productivity gap followed a similar course.22
FIGURE 5.2. Japan’s Catch-up to the United States in Productivity Stalls in the 1990s. Source: Jorgenson, Nomura, and Samuels 2018, 18.
Key Technological Changes in the IR-3
Parsing through the different trajectories of technological change is a necessary first step to determine whether the LS and GPT mechanisms were operative in this period. This task is complicated by the tremendous technological changes that emerged in the IR-3, such as the first microprocessor (1971), the production of recombinant DNA (1972), the VHS format for video recording (1976), and the first personal computer (1981). Guided by past scholars’ efforts to map the key nodes of the information revolution as well as analytic measures for leading sectors and GPTs, this section takes stock of key technological drivers that affected the US-Japan economic power balance.
Candidate Leading Sectors
Amid a shifting technological landscape, the most likely sources of LS trajectories were information and communications technologies (ICTs). Certainly, scholars have highlighted technological developments in a wide range of industries as possible leading sectors, including lasers and robotics.23 Nonetheless, Japan’s success in the computer, consumer electronics, and semiconductor industries was most relevant for its prospects of overtaking the United States as the foremost economic power. In each of these three leading sectors, Japan dominated the production of key components.24
All three relatively new industries achieved extraordinary growth off the back of technological breakthroughs, fulfilling the established criteria for leading sectors. In the US economy during the 1980s, computer and data-processing services ranked as the fastest-growing industries in terms of jobs added.25 After Japan’s MITI identified semiconductors and computers as strategic industries in 1971, both industries experienced extremely high growth rates in the next two decades.26 The US electronics industry also experienced a remarkable surge during the late twentieth century, growing thirty times faster than other manufacturing industries by some estimates.27 These trends in computers and electronics held across advanced industrialized countries.28
Candidate GPTs
Given that the IR-3 is often known as the “information revolution,” clusters of ICT innovations naturally serve as the most likely sources of GPT trajectories. Efforts to map this era’s GPTs specifically highlight computers,29 semiconductors,30 and the internet.31 Each of these technological domains exhibited great scope for improvement and complementarities with other technologies.
Because advances in computers, semiconductors, and the internet were all closely connected, I group technological developments in these three domains under “computerization,” the process in which computers take over tasks such as the storage and management of information. This is consistent with other studies that identify the general category of ICT as the GPT operative in this period.32 The growing prevalence of software-intensive systems enabled computers to become more general-purpose. Computerization also benefited from advances in both semiconductors, which reduced costs for investments in IT equipment, and the internet, which connected computers in efficient networks.33
Sources of LS and GPT Trajectories
In sum, the IR-3’s candidate leading sectors and GPTs all revolved around ICTs (table 5.1). Other technical advances in lasers, new sources of energy, and biotechnology also drew attention as possible sources of LS and GPT trajectories. I do not trace developments in these technologies because their potential was largely unrealized, at least within the context of US-Japan economic competition during the IR-3.
Though candidate leading sectors and GPTs converge on ICTs, they diverge on the key trajectories. The GPT perspective emphasizes the process by which firms transfer tasks and activities to computers. In contrast, LS accounts spotlight the growth of key industry verticals. For example, the consumer electronics industry fits the mold of previous candidate leading sectors like automobiles and cotton textiles, which were large and fast-growing but limited in their linkages to other industries. Taking stock of candidate leading sectors and GPTs merely functions as a preliminary filter. The rest of the chapter will further flesh out the differences between the LS- and GPT-based explanations of how these technologies affected the US-Japan economic rivalry.
table 5.1 Key Sources of Technological Trajectories in the IR-3
Candidate Leading Sectors
Candidate GPTs
Computer industry
Computerization
Consumer electronics industry
Semiconductor industry
GPT vs. LS Mechanisms: The (Non) Spread of ICTs across Japan’s Economy
In both the IR-1 and IR-2 cases, a technological revolution sparked a shift in global economic leadership. The case analyses aimed to determine whether the historical evidence fit better with observable implications derived from the GPT or LS mechanism. Revolutionary technological breakthroughs also occurred in the IR-3, but an economic power transition never occurred. Thus, if the case study reveals that Japan dominated innovation in the computer, consumer electronics, and semiconductor industries, then this would provide disconfirming evidence against the LS mechanism. Likewise, if the historical evidence shows that Japan led the way in computerization during this period, this would undermine GPT diffusion theory.
LS Mechanism Present: Japan Dominates the Production of Key ICTs
A bevy of evidence establishes that the LS mechanism was operative during the IR-3. From the mid-twentieth century through the 1980s, Japan captured a growing global market in new industries tied to major technological discoveries in semiconductors, consumer electronics, and computers. In dynamic random-access memory (DRAM) chips, one of the highest-volume verticals in the semiconductor industry, Japanese firms controlled 76 percent of the global market share.34 By one US federal interagency working group estimate, from 1980 to 1987 the United States lost the lead to Japan in more than 75 percent of critical semiconductor technologies.35
Japanese industry also gained competitive advantages in consumer electronics. From 1984 to 1990, US firms lost global market share in thirty-five of thirty-seven electronics categories as Japanese firms took over the production of many electronic products.36 Japan occupied dominant shares of global production of color televisions and DVDs.37 It was also the first economy to commercialize high-definition television (HDTV) systems, a highly touted part of the consumer electronics market.38
A similar trend held in computers, especially in computer hardware components like flat panel displays.39 The US trade balance in computers with Japan turned from a surplus in 1980 into a $6 billion deficit by 1988.40According to the Yearbook of World Electronics Data, in 1990 Japan’s share of global computer production eclipsed the share held by the United States, which had previously led the world.41
Comparisons of LS growth rates also indicate that Japan was poised to overtake the United States as the economic leader. In an International Organization article published in 1990, Thompson posited that average annual growth rates in leading sectors across major economies heralded shifts in economic leadership. Over the nineteenth century, Britain’s growth rate in leading sectors peaked in the 1830s before flattening between 1860 and 1890, a period when the United States and Germany outstripped Britain in LS growth rates.42 Crucially, Thompson’s data showed that Japan outpaced the United States in growth rates within leading sectors from 1960 to 1990.43 Linking these historical trends, Thompson identified Japan as America’s main competitor for “systemic leadership.”44
Comprehensive assessments of Japan’s relative industrial strength support this account of LS growth rates. US government, academic, and industry entities issued a plethora of reports warning of Japan’s growing global market share and exports in key technologies. One review of six such reports, all published between 1987 and 1991, found a growing consensus that US capabilities in many of these technologies were declining relative to Japan’s.45 A 1990 US Department of Commerce report on trends in twelve emerging technologies, including supercomputers, advanced semiconductor devices, and digital imaging technology, projected that the United States would lag behind Japan in most of these technologies before 2000.46 The 1989 MIT Commission on Industrial Productivity’s Made in America: Regaining the Productive Edge serves as a particularly useful barometer of Japan’s position in leading sectors.47Made in America argued that the United States was losing out to Japan in eight manufacturing sectors, including consumer electronics, semiconductors, and computers. As business historian Richard Langlois summarizes, “By the mid-1980s, by most accounts, America had ‘lost’ consumer electronics and was in imminent danger of losing semiconductors and computers.”48
Some argued that Japan’s advantage in these leading sectors was rooted in certain institutional arrangements. Observers regularly pointed to Japan’s keiretsu system, which was structured around large, integrated business groups, as the key institutional factor in its success in high-tech industries. The MIT Commission’s Made in America report, for instance, questioned whether the US system of industrial organization could match up against “much stronger and better organized Japanese competition.”49 This aligned with a common narrative in the mid-1980s that “American firms should become more like Japanese firms.”50
Others pointed to Japan’s industrial policy, coordinated by MITI, as the key institutional competency that explained Japan’s success in leading sectors. Academics and policymakers pushed for the United States to imitate Japan’s industrial policy approach, which they perceived as effective because of MITI’s ability to strategically coordinate R&D investments in key technologies.51 For instance, scholars regarded the “Fifth Generation Project,” a national initiative launched by MITI in 1982, as a stepping-stone to Japan’s building of the world’s most advanced computers.52 The American aversion to industrial policy and a decentralized economic policymaking apparatus, by comparison, was alleged to be detrimental to innovation in the IR-3’s new technologies.
By the turn of the millennium, such arguments were no longer being put forward. Despite capturing key LS industries and rapidly catching up to the United States in the 1980s, Japan did not ultimately overtake the United States as the lead economy. Contrary to the expected outcome of the LS mechanism, Japan’s control of critical sectors in semiconductors and consumer electronics did not translate into strong, sustained economic growth. This outcome challenges the LS mechanism’s validity in the IR-3 case.
An Absent GPT Mechanism: The United States Leads Japan in ICT Diffusion
Does evidence from the IR-3 also discredit the GPT mechanism? If the components of the GPT mechanism, like the LS mechanism, were present during this period, this would weaken the explanatory power of GPT diffusion theory. However, in contrast to its success in key leading sectors, Japan lagged in adopting computerized technologies. Thus, GPT diffusion theory would not expect Japan to have overtaken the United States in the IR-3.
To account for sustained US economic leadership in the IR-3, this section traces the developments of the IR-3 in the United States and Japan across the three dimensions that differentiate GPT from LS trajectories. First, relative to the LS mechanism, the impact timeframes of the IR-3’s technological breakthroughs are more elongated. Developments in ICTs did not spread to a wide range of economic applications until the 1990s. Second, though Japan excelled in the production of computers and electronics, it fell behind in the general pace of computerization across the economy. Lastly, Japan’s advantages were concentrated in a narrow range of ICT-producing industries, whereas the United States benefited from broad-based productivity growth.
IMPACT TIMEFRAME
The advance of computerization, like past GPT trajectories, demanded a prolonged period of organizational adaptation and complementary innovations. It is reasonable to date the computerization GPT’s emergence as the year 1971—the year when Intel introduced the microprocessor, which greatly expanded the functionalities of computers.53 It was also the year when the share of information technology equipment and software reached 1 percent in the net capital stocks of the median sector in the US economy.54 Before then, during the 1960s, mainframe computers powered by integrated circuits serviced only a limited range of commercial purposes, such as producing bank statements and managing airline reservations. With the internet’s rise in the 1990s, new information and communication networks further spread computerization to different business models, such as e-commerce.55 Alongside this stream of complementary technical advances, firms needed time to build up their computer capital stock and reorganize their business processes to match the needs of information technology.56
Computers traveled a gradual and slow road to widespread use. By the late 1980s, many observers bemoaned the computer revolution’s failure to induce a surge of productivity growth. In 1987, the renowned economist Robert Solow distilled this “productivity paradox” in a famous quip: “We see the computers everywhere but in the productivity statistics.”57 A decade later, however, the growing adoption of information technology triggered a remarkable surge in US productivity growth.58 It took some time, but the American economy did eventually see the computers in the productivity statistics.
The landscape of US-Japan technology competition looks very different when accounting for the elongated lag from computerization’s arrival to its widespread diffusion. Japan’s control over key segments of ICT production in the 1970s and 1980s did not correspond to an advantage in GPT diffusion. A more patient outlook illuminates that the delayed impact of computerization aligns with when the United States extended its productivity lead over Japan. After 1995, while Japan’s economic rise stalled, labor and total factor productivity grew rapidly for a decade in the United States. The difference was that the United States benefited greatly from an ICT-driven productivity acceleration.59
FIGURE 5.3. The US-Japan Computerization Gap Widens in the 1990s. Source: Milner and Solstad 2021; Comin 2010, 381.
PHASE OF RELATIVE ADVANTAGE
If the GPT mechanism was operative for Japan’s rise in the IR-3, Japan should have led the United States in computerization. Though Japan continued to contest US leadership in the production of certain computer architectures and devices, it failed to keep up with the United States in the adoption of computers across industries. As figure 5.3 reveals, the gap between the United States and Japan in computerization widened in the 1990s. In fact, South Korea, which lagged behind Japan in generating new innovations in computer systems, surpassed Japan in its computer usage rate during the 1990s. Taken together, these indicators suggest that Japan’s problem was with GPT diffusion, not innovation.
The pathway by which ICTs drove the US-Japan productivity gap is particularly revealing. In sectors that produced ICTs, Japan’s TFP acceleration was similar to the US trajectory; however, in sectors that intensively used IT, Japan’s TFP growth lagged far behind that of its rival.60 In particular, US ICT-using service industries adapted better to computerization. In terms of labor productivity growth in these industries, the United States experienced the strongest improvement out of all OECD countries from the first half of the 1990s to the second half of the decade.61 The contribution of ICT-using services to Japan’s labor productivity growth, by contrast, declined from the first half to the second half of the decade.62
Japan eventually adopted the GPT trajectory associated with ICT. Like all advanced economies at the frontier, Japan could draw from the same technology pool as America. Using a growth accounting framework that accounts for cyclical factors, Takuji Fueki and Takuji Kawamoto trace Japan’s post-2000 resurgence in TFP growth to the extension of the ICT revolution to a broader range of IT-using sectors.63 By then, however, Japan was at least five years behind the United States in taking advantage of computerization.
BREADTH OF GROWTH
Alongside the dispersion of ICTs throughout the US economy, the sources of productivity growth also spread out. In the United States, spillovers from ICT advances, especially in services, contributed to economy-wide TFP growth. Industry-level patterns of TFP growth reveal that US productivity growth became noticeably more broad-based after 1995, a trend that accelerated further after 2000.64
In contrast, Japan’s advantages in the IR-3 were concentrated in a narrow range of sectors. After 1995, Japanese productivity growth remained localized, only transitioning to a more broad-based pattern after 2000.65 Michael Porter’s exhaustive account of national competitiveness across leading economies describes Japan as a “study in contrasts,” with some of the most internationally competitive industries found side by side with some of the most uncompetitive.66 The hype over Japan’s success in leading sectors induced some analysts to generalize from a few exceptional industrial sectors while overlooking developments in struggling sectors.67
LS Mechanisms and “Wintelism”
Some international political economy scholars divide trends in US competitiveness into a phase of relative decline in the leading sectors of the 1980s (consumer electronics, computer hardware, and parts of the semiconductor industry) and a period of resurgence in the new leading sector of the 1990s (software electronics).68 This explanation for why Japan’s LS advantage did not convert into an economic power transition could conceivably restore credibility to the LS mechanism. However, even this generous interpretation fails to capture the dynamics of the IR-3.69
Consider one prominent line of LS-based thinking that emphasizes US advantages in adapting to “Wintelism,” a type of industrial structure best suited for new advances in software electronics. Wintelism, a portmanteau of “Windows” and “Intel,” refers to the transformation of the computer industry from a vertically integrated oligopoly into a horizontally segmented structure dominated by components providers with controlling architectural standards, such as Intel and Microsoft.70 Compared to Japan, the US institutional environment was more supportive of horizontal, specialized value-chains in software electronics. It is put forward that Japan’s inability to adapt to the Wintelist industrial paradigm explains why it was unable to overtake the United States as the leading economy.71
The Wintelism argument falls prey to general issues with the LS mechanism.72 Like LS accounts of Japan’s advantage in semiconductors and consumer electronics in the 1980s, the Wintelism argument still places too much value on ICT-producing industries. Capturing profit shares in the global software electronics industry is not the same as translating new advances in software electronics into economy-wide growth.73 The pathway from new technologies to overall productivity growth involves much more than just the success of companies like Microsoft and Intel.
Indeed, as a GPT diffuses more widely, large monopolists may hinder coordination between the GPT and application sectors. Both Microsoft and Intel, for instance, often restricted sharing information about their technology roadmaps, thereby hindering complementary innovations and adoption of microelectronics in applications sectors such as automobiles. In fact, regulatory and technological forces were necessary to limit the influence of dominant firms and encourage the development of complementary technologies, which were crucial to widening the GPT trajectory of computerization.74 It is conceivable that the United States could have achieved a greater rate of computerization if its computer industry had not been dominated by two firms that held key architectural standards.
Overall, hindsight is not kind to LS-based accounts of Japan’s institutional advantages in the IR-3. While matching institutional competencies to particularly significant technological trajectories is a sound approach, the LS trajectory fails to capture how technological changes opened opportunities for an economic power transition. Japan’s industrial structure and sectoral targeting policies reaped economic gains that were temporary and limited to specific industries.75 To understand why the United States gained lasting and broad-based advantages from computerization, a different set of institutional complementarities must be explored.
Institutional Complementarities: GPT Skill Infrastructure in the IR-3
In line with GPT diffusion theory, institutional adaptations that widened the base of computer engineering skills and knowledge proved crucial to the enduring technological leadership of the United States in the IR-3.76 Computerization required not just innovators who created new software architectures but also programmers who undertook more routine software engineering tasks. It was the US ability to tap into a more expansive pool of the latter that fueled a more intensive pace of computerization than in Japan.
Widening the Base of Computer Engineers
GPTs generate an imbalance between the possibility of sweeping changes across many domains and the constraints of existing skills. Historically, engineering disciplines have developed procedures for adapting new GPT-linked knowledge across localized applications and expanded access to such knowledge to a broader set of individuals. Similarly, the key element of the American GPT skill infrastructure in the IR-3 was the development of a computer science discipline—the latest in a line of engineering disciplines that have emerged in the wake of GPTs.77
US education effectively adapted to changes in the computerization trajectory. The recognition of computer science as an independent discipline, as evidenced by the early and rapid growth of computer science departments in the United States, helped systematize the knowledge necessary for the spread of computerization.78 Led by top universities and the Association of Computing Machinery (ACM), US institutions piloted new training programs in computer science. In 1968, the ACM published an influential and forward-looking curriculum that helped colleges organize their computing education.79 These adaptations converted the practical experience from developers of new computer and software breakthroughs into general, accessible knowledge.80
The development of this GPT skill infrastructure met significant hurdles. In the 1970s, the US computer science discipline struggled to meet demands for software engineering education in the midst of conflicts over the right balance between theory-oriented learning and hands-on programming work.81 Reacting to this situation, the ACM’s 1978 curriculum revision directed computer science toward application-based work, stating that programming exercises “provide a philosophy of discipline which pervades all of the course work.”82 Industry pressure and the US Department of Defense’s Software Engineering Institute, a partnership established with Carnegie Mellon University in 1984, expanded the number of software engineering specializations at universities.83 Teaching capacity was another inhibiting factor. Computer science departments had to limit course enrollments because they could not fill the faculty positions to meet exploding student demand, resulting in a decline in computer science enrollments in the mid-1980s.84
Though the process was not seamless, overall trends reflect the US success in widening the pool of engineering skills and knowledge for advancing computerization. From 1966 to 1996, the number of undergraduate computer science degrees awarded annually in the United States grew from 89 to about 24,500.85 In 1986, at its peak during this period, computer science accounted for 12.5 percent of all science and engineering degrees awarded in the United States.86 According to benchmarking efforts by the Working Group for Software Engineering Education and Training, US institutions accounted for about one-third of the world’s bachelor’s degree programs in software engineering in 2003.87 Throughout this period, the United States also benefited from a system open to tapping foreign software engineering talent.88
The US Comparative Advantage over Japan in GPT Skill Infrastructure
Did GPT skill infrastructure factor decisively in Japan’s inability to keep pace with the United States in the diffusion of ICTs? Varying data reporting conventions, especially the fact that Japanese universities subsumed computer science under broader engineering fields, make it difficult to precisely quantify the US-Japan gap in software engineering skills.89 One narrative that gained momentum in the late 1980s accredited Japan’s success in high-tech manufacturing industries to its quantitative advantage in engineers. A National Science Foundation (NSF) report and two State of the Union addresses by President Ronald Reagan endorsed this belief.90 For example, the NSF’s 1997 special report on Japan’s scientific and technological capabilities declared, “By 1994, with roughly one-half the population, Japan produced more engineering and computer science degrees at the undergraduate level than the United States.”91 Such statements blended computer science degrees with all categories of engineering education.
Computer science–specific data reveal the US edge in ICT human resources. According to data from Japan’s Information Technology Promotion Agency, Japan awarded about 16,300 computer science and mathematics bachelor’s degrees in 2009, while the United States awarded 63,300 of these types of degrees that same year.92 One survey by the Japanese Information Technology Services Industry Association found that only 3.6 percent of college graduates who entered Japan’s information service industry in April 1990 received their degree from a computer science department.93 By counting immigrants entering computer-related professions and total university graduates in ICT software and hardware fields, one study estimated annual inflows into the US and Japanese ICT labor pools. In 1995, these inflows into the US ICT labor pool outpaced those in Japan by 68 percent. By 2001, this gap between the two countries’ annual inflows of ICT talent had reached almost 300 percent.94 Therefore, in the years when the US advantage in ICT diffusion was most pertinent, the skill gap between the United States and Japan in computer and software engineering talent grew even wider.95
Moreover, a computer science degree in Japan did not provide the same training as one in America. First, Japanese universities were slow to adapt to emerging trends in computer science. In both 1997 and 2007, the Information Processing Society of Japan modeled its computing curriculum revisions on American efforts that had been made six years earlier.96 The University of Tokyo, Japan’s leading university, did not establish a separate department of computer science until 1991, which was twenty-six years later than Stanford.97 Overly centralized governance of universities also inhibited the development of computer science as an independent discipline in Japan.98 As Jeffrey Hart and Sangbae Kim concluded in 2002, “The organizational and disciplinary flexibility of US universities in computer science has not been matched in any of the competing economies.”99
Software engineering presented a particular challenge for Japan. In 1988, the Japanese-language industry journal Nikkei Computer surveyed six thousand Japanese firms that used office computers. Situated outside the computer industry, these firms were involved in a broad range of fields, including materials manufacturing, finance, services, government, and education. More than 80 percent of the responding companies disclosed shortages of software programmers and designers.100 On average, outside firms provided one-quarter of their information technology personnel, and their reliance on outsourcing was magnified for programmers, system designers, and application managers.101 A nonprofit foundation for ICT development in Japan reported similar barriers to computer utilization in 1991. The survey found that companies relied heavily on computer personnel, especially software engineers, dispatched temporarily from other organizations.102 Small and medium-sized software departments, which could not afford to invest in on-the-job training, were especially disadvantaged by the lack of formal software engineering education in Japan.103
Bibliometric techniques can help substantiate the gap between the United States and Japan in skill infrastructure for software engineering. I analyzed around seven thousand publications from 1995 in the Web of Science Core Collection’s “Computer Science, Software Engineering” category.104 To gauge the breadth of institutional training in software engineering, I counted the number of Japanese and American universities that employed at least one researcher represented in this dataset. According to my estimates, the United States boasted 1.59 universities per million people that met this baseline quality of software engineering education, while Japan only had 1.17 universities per million people. This amounts to a gap of around 40 percent.
Lastly, weak industry-university linkages in computer science hampered Japan’s development of GPT skill infrastructure. Imposing centralized control over universities, the Japanese Ministry of Education, Science, and Culture (MESC) inhibited cooperation between new departments of information science and the corporate labs where much of the computing talent was concentrated.105 Japanese researchers regularly complained about the size of MESC grants, as well as the ministry’s restrictions on their ability to seek alternative funding sources. Japan’s overall budget level for university facilities in 1992 remained the same as it was in 1975. Additional government funds went instead to independent centers of excellence, diverting resources away from efforts to broaden the pool of training institutions in software engineering.106
Alternative Factors
How do alternative explanations perform in this case? A range of factors beyond GPT skill infrastructure could have influenced diverging rates of ICT diffusion in the United States and Japan. I focus on the role of external threats and varieties of capitalism because they present alternative mechanisms for how states adapted differently to the technological revolution that occurred in the IR-3.
Threat-Based Explanations
Threat-based theories struggle to account for differences in the US and Japanese technological performance in this period. A “cult of vulnerability” permeated Japan’s leaders over this period as they coped with tensions in East Asia and the oil crises of the 1970s.107 Likewise, the growth of the US “national security state,” fueled by the dangers of the Cold War, functioned as “the secret to American innovation.”108 Under his creative insecurity framework, Taylor holds up both Japan and the United States as exemplars of the IR-3 period, reasoning that they both partly owed their technological success to the galvanizing effects of a threatening international environment.109 General threat-based explanations therefore cannot explain differences in technological outcomes between the United States and Japan, namely, why the United States was more successful in ICTs than Japan.
A related argument points to the significance of US military procurement for computerization. As was the case with its influence on the American system of manufacturing in the IR-2, the US military provided the demand for initial investments in computers and semiconductors. In the 1940s and 1950s the US military was a key patron behind computing breakthroughs.110 Assured by large military procurements, innovative firms undertook risky, fundamental research that produced spillovers to many other industries. For instance, the first all-purpose electronic digital computer, the University of Pennsylvania’s electronic numerical integrator and calculator (ENIAC), was developed during World War II. The ENIAC was supported by funding from the Army Ballistics Research Laboratory, and the first program run on the computer was a simulation of the ignition of the hydrogen bomb.111
In place of the military, could other entities have served as a large demand source for ICTs? Commercial entities like Bell Labs and IBM also developed fundamental breakthroughs in ICTs. According to Timothy Bresnahan and Manuel Trajtenberg, it was “only a coincidence” that US government demand played such a pivotal role in semiconductor development.112 Others argue that while commercial advances in semiconductors and computers would likely still have occurred absent the impetus of military funding, they would have emerged after substantial delay.113
Resolving this debate depends on one’s view of the key stage in computerization. Those who emphasize the importance of military procurement often hold up the importance of first-mover advantages in the American computer industry.114 However, decades after the military helped develop the first computers and transistors, Japan had cornered the market in many related industries. The significance of military procurement is diminished when a GPT’s dissemination, as opposed to its emergence, is taken as the starting point. By 1960, the start of the IR-3 period, ICT development in the United States was already much less reliant on military support. In 1955, the demand for Bell’s transistors from two large telephone networks alone was nearly ten times more than from all military projects.115 In fact, as the commercial sector increasingly drove momentum in ICTs, military involvement arguably hindered continued advances in the commercial sector, as there was tension between the different technical cultures.116
On balance, the most significant aspect of the military’s involvement in the advance of computerization in the United States was its role in building up GPT skill infrastructure. The US military played a key role in cultivating the computer science discipline in its early years. Beginning in the 1960s, defense agencies supported academic research in computer science, such as the aforementioned Software Engineering Institute, which created centers of excellence and broadened the base of computer science education.117 From 1977 through the mid-1980s, defense funding supported more than half of academic computer science R&D.118 At the same time, military investment in computer science did not come without downsides. Defense funding was concentrated in elite research universities at the cutting edge of the field, such as Carnegie Mellon and Stanford, whereas nondefense government funding supported computer science education across a wider range of US universities.119 On the effects of military computer science funding, Stanford professor Terry Winograd wrote, “It has resulted in a highly unequal situation in which a few schools have received almost all the resources. Although this may have led to more effective research in the short run, it has also been a factor contributing to the significant long-term shortage of trained computer researchers.”120
VoC Explanations
The varieties-of-capitalism (VoC) approach provides another possible explanation for why the US economy benefited more than Japan’s from the innovations of the IR-3.121 According to the VoC framework, firms in coordinated market economies (CMEs) provide industry-specific training that is more conducive to incremental innovation, whereas worker training in more general skills in liberal market economies (LMEs) proves more favorable for radical innovations. VoC scholars point to some evidence from the international pattern of innovation during the IR-3 that supports these expectations. Based on patent data from 1983–1984 and 1993–1994, Peter Hall and David Soskice find that Germany, a CME, specialized in technology classes characterized by incremental innovation, whereas the United States, an LME, specialized in domains characterized by radical innovation.122 Therefore, the VoC perspective expects that Japan, a CME like Germany, was unable to keep up with the United States in the IR-3 because high-tech sectors such as computer software and biotechnology demanded radical innovations.123
This VoC-derived explanation provides an incomplete account of the IR-3 case. First, comprehensive empirical investigations into the innovative performance of CMEs and LMEs, especially the success of Japan as a radical innovator, undermine the explanatory power of VoC theory for the IR-3 period. Hall and Soskice’s initial analysis was based on four years of data on patent counts from only two countries, the United States and Germany. Taylor’s more extensive analysis, which covered thirty-six years (1963–1999) of patent counts and forward citations for all LME and CME countries, found that the predictions of VoC theory are not supported by the empirical data.124 In fact, contrary to the expectations of the VoC explanation, Japan was a leader in radical innovation, ranking second only to the United States in patent counts weighted by forward citations, which are a strong proxy for the radicalness of innovations.125
Second, VoC theory does not make distinctions between different types of general skills, which varied in their significance to national success in the IR-3. Regarding general training in terms of foundational schooling, Japan was making substantial improvements in average years of schooling, enrollment ratio, and access to higher education.126 GPT diffusion theory specifies the key general skills as those that best suited the advance of computerization. Consistent with these expectations, the case study evidence points to the US-Japan gap in software engineering, a set of general skills that permeated sectoral boundaries, as the crucial factor in US success with widespread computerization.
Case-Specific Factors
Other factors unique to the IR-3 case deserve further consideration. Among these alternative explanations, one popular theory was that Japan’s kanji system (Chinese-based characters) contributed to its slow adoption of computers.127 Marshall Unger, a specialist in the Japanese writing system, highlighted difficulties with representing kanji in computerized formats, which resulted in higher costs for storing data and for word-processing functions.128 American computers had to handle only ninety-five printable characters, whereas Japanese personal computers needed to store six thousand Japanese characters.129 Not only did language differences increase the cost of Japanese computers, but they also prevented Japanese adopters from using off-the-shelf computers from the United States, as these did not support Japanese language functions.
While particularities of the Japanese language may have initially hindered Japan’s computerization, it is important not to overstate the impact of this language barrier. In a review of this theory, another expert on computational linguistics argued that Unger overemphasized the additional overhead and speed costs associated with Japanese writing systems.130 Moreover, users and companies adapted over time. By the end of the 1980s, advances in processor technology allowed computers to support the greater word-processing demands of Japanese language systems.131 Therefore, during the critical years when the US-Japan computerization gap widened, the impact of the kanji system was less pronounced.
Summary
Through much of the late twentieth century, it was only a matter of time until Japan achieved economic preeminence—at least in the eyes of many analysts and scholars. Invoking the assumptions of the LS mechanism, they expected that this economic power transition would be brought about by Japan’s advantages in new sectors such as consumer electronics, semiconductor components, and computer hardware. Today, after Japan’s decade-long slowdown in productivity growth, there is virtually no discussion of it overtaking the United States as the leading economic power.
Looking back, one might be tempted to conclude that history has vindicated past critics who labeled the claims of Japan’s imminent ascension to technological hegemony as “impressionistic,”132 as well as the retrospective analyses that called out such projections for being “premature.”133
This chapter’s conclusions suggest a more nuanced interpretation. It is not that the prognoses of LS-based accounts were overeager or overly subjective. The real issue is that they were based on faulty assumptions about the pathway by which technological advances make economic power transitions possible. Indeed, the IR-3 case provides strong negative evidence against the LS mechanism, revealing that the expected outcome of an economic power transition failed to materialize in part because of the US advantage in GPT diffusion. The relative success of the United States in diffusing the trajectory of computerization across many ICT-using sectors, in line with GPT diffusion theory, was due to institutional adaptations to widen the skill base in computer engineering. In sum, the US advantage in GPT diffusion accounts for why the economic power transition expected by the LS account failed to transpire.
6 A Statistical Analysis of Software Engineering Skill Infrastructure and Computerization
I HAVE ARGUED that the shape of technological change is an overlooked dimension of the rise and fall of great powers. Most researchers point to various institutions to explain why some countries experience more scientific and technological progress than others. A central insight of this book is that the institutional factors most relevant for technological leadership depend on whether key technological trajectories map onto GPT diffusion or LS product cycles. GPT diffusion theory posits that great powers with better GPT skill infrastructure, defined as the ability to broaden the engineering skills and knowledge linked to a GPT, will more effectively adapt to technological revolutions.
This chapter evaluates a key observable implication of GPT diffusion theory. The expectation is that where there is a wider pool of institutions that can train engineering talent related to a GPT, there will be more intensive rates of GPT diffusion. Using data on computerization and a novel approach to estimate the number of universities that provide quality software engineering education in a country, this chapter first tests the theorized connection between GPT skill infrastructure and GPT adoption on time-series cross-sectional data for 19 advanced and emerging economies from 1995 to 2020. I supplement this panel analysis with two additional tests: a duration model of the speed by which 76 countries achieved a certain computerization threshold, as well as a cross-sectional regression of data on 127 countries averaged over the 1995–2020 period. While the historical case studies verified this relationship among great powers, large-n quantitative analysis allows us to explore how GPT diffusion applies beyond the chosen case studies.
To preview the findings, the evidence in this chapter backs GPT diffusion theory. Countries better positioned to widen their base of software engineering skills preside over higher rates of computer adoption. This relationship holds even when accounting for other factors that could affect computerization and different specifications of the independent and dependent variables. This chapter proceeds by operationalizing computerization rates and skill infrastructure in software engineering and then statistically testing the relationship between the two variables.
Operationalizing the Independent Variable: GPT Skill Infrastructure in Software Engineering
My key independent variable is skill infrastructure connected to computerization. The computer, a prototypical GPT, represents a natural choice for this type of inquiry, as engineering education data for many past GPTs is nonexistent for many countries.1 Plus, enough time has passed for us to see the effects of computerization. The statistical analysis focuses on the effects of skill formation institutions in software engineering, the computer science discipline tasked with training generalists in computing technology.2 Concretely, this chapter’s measure of GPT skill infrastructure captures the breadth of a country’s pool of software engineering skills and knowledge.
Efforts to measure the GPT skill infrastructure in software engineering face three challenges. First, standardized measures of human capital in computer science across countries are not available. The UNESCO Institute for Statistics (UIS) collects internationally comparable data on technicians and researchers in various fields, but this dataset does not include information specific to computer science and has limited temporal coverage.3 Second, variation across countries in the format of computer science education undercuts some potential benchmarks, such as undergraduate enrollments in computer science programs. In some countries, computer science education is subsumed under a broad engineering category, not recognized as a separate degree course.4 Lastly, comparisons of computer science education struggle to account for the quality of such training. International rankings of universities for computer science garner media coverage, but they rely on subjective survey responses about reputation and largely concentrate on elite programs.5
To the extent possible, my measure of GPT skill infrastructure addresses these obstacles. The goal is to operationalize engineering-oriented computer science education in a way that can be standardized across countries and accounts for differences in the format and quality of computer science education. My novel approach estimates the number of universities in each country that can be reasonably expected to provide a baseline quality of software engineering education. To establish this baseline in each country, I count the number of universities that employ at least one researcher who has published in a venue indexed by the Web of Science (WoS) Core Collection’s category on “Computer Science, Software Engineering.” In this category, the WoS citation database extends back to 1954 and allows for reliable cross-country comparisons based on institutional affiliations for published papers and conference proceedings.6 This approach is also insulated from distinctions related to whether certain degrees count as “computer science” programs or as “general engineering” courses. A particular university’s naming scheme has no bearing; as long as an institution retains at least one researcher who has published in the software engineering field, it counts in the GPT skill infrastructure measure.
To gather data on the number of universities that contribute to software engineering skill formation around the world, I analyze 467,198 papers from the WoS Core Collection’s “Computer Science, Software Engineering” category published between the years 1995 and 2020. I use the Bibliometrix open-source software to derive institutional and country affiliations from this corpus.7 Specifically, I collect the university and country affiliations for the corresponding authors of all 467,198 publications. For each country, I count the number of distinct university affiliations. Hypothetically, if country X’s researchers were all concentrated at a single center of excellence, it could boast more researchers represented in the corpus than country Y but still score lower on my metric. For making comparisons across countries, the number of distinct university affiliations is a better indicator of a country’s access to a broad pool of training institutions for software engineering, which is central to GPT skill infrastructure.
FIGURE 6.1. Software Engineering Skill Infrastructure by Country (2007). Source: Author’s calculations based on Web of Science Core Collection database.
I estimate a country’s GPT skill infrastructure in a particular year by averaging its score on this metric in that year along with its scores in the two previous years. This step provides checks against the risk that developments specific to a particular year—a conference cancellation, for example—may muddle the measure. To illustrate country distributions on this metric, figure 6.1 depicts the number of universities that meet my baseline for software engineering skill formation in G20 countries for one of the middle years in the dataset.
As with all bibliometric research, given the bias toward English-language papers in publication datasets, this method could undercount publications from non-English-speaking countries.8 Fortunately, this linguistic bias found in social science papers is less pronounced in engineering and mathematics papers, which comprise my dataset.9 Another factor that mitigates this bias is the very low standard for quality engineering education. Even if an institution’s researchers publish a substantial portion of their writings in a non-English language, as long as just one publication landed in the WoS “Computer Science, Software Engineering” category, that institution would still count in my definition of GPT skill infrastructure.
I considered other measures of software engineering skills, but none were suitable for this type of analysis. Data on the number of full-time equivalent telecommunication employees, collected by the ITU, shed some light on the distribution of ICT skills in various economies. Concretely, this indicator captures the total number of people employed by telecommunication operators for the provision of fixed-telephone, mobile-cellular, and internet and data services.10 The rationale is that the number of employees in this critical ICT services industry represents the broader pool of software engineering talent in a country. However, since this measure is biased toward specialists who develop and install computer technologies and other ICTs, it overlooks many engineers who intensively use ICTs in their work, even if they are not involved in developing software and computing tools.11
The time coverage of other measures was limited. For instance, the International Telecommunication Union (ITU) database on ICT skills like programming or coding in digital environments only goes back to 2019.12 In its Global Competitiveness Index, the World Economic Forum surveys business executives on the digital skills of their country’s population, but this data series starts in 2017.13
Operationalizing the Dependent Variable: Computerization Rates
Divergences among nations in scientific and technological capabilities have attracted a wide range of scholarship. While my focus is on the overall adoption rate of GPTs within economies, many scholars and government bodies have made significant contributions to quantifying national rates of innovation, often based on patenting activity, publications, and R&D investments.14 Cross-national data on the diffusion of specific technologies, by comparison, has been sparse.15 The Cross-country Historical Adoption of Technology (CHAT) dataset, which documents the intensity with which countries around the world use fifteen historically significant technologies, has helped address this deficiency.16 Other studies of cross-national technology adoption gaps quantify the diffusion of the internet and government uses of information technology.17
My primary measure of computerization is the proportion of households with a computer. These data are sourced from the International Telecommunication Union’s World Telecommunication/ICT Indicators (WTI) database.18 In this dataset, access to a computer includes use of both desktop and portable computers but excludes devices with some computing ability, such as TV sets and mobile phones.19 By estimating the number of households in a country with access to a computer, this measure elucidates cross-country differences in the intensity of computerization. Though observations for some countries start in 1984, there is limited coverage before 1995, which serves as the initial year for the data collection effort detailed in this chapter.
After the ITU was tasked with supplying indicators for access to ICTs around the world—a crucial target of the United Nations’ Millennium Development Goals (MDG) adopted in 2000—the agency started to track the number of personal computers by country.20 The ITU produces computer usage figures through two methods. First, when available, survey data from national and supranational statistical offices (such as Eurostat) are used. Though the MDG initiative has encouraged national statistical offices to help the ITU in monitoring ICT access, data coverage is still incomplete. If data on the number of households with a computer are unavailable for a country in one year, the ITU makes an estimate based on computer sales and import figures, adjusted to incorporate the average life of a computer as well as other related indicators, such as the number of computer users. For example, the computer usage indicator for Latvia comes from Eurostat in 2013, an ITU estimate in 2014, and the Central Statistical Bureau of Latvia in 2015.
Despite its limitations, I prefer the ITU’s computerization indicator over alternative measures. Francesco Caselli and Wilbur John Coleman examine the determinants of computer adoption across a large sample of countries between 1970 and 1990. To estimate the intensive margin of diffusion, they use the value of a country’s computing equipment imports per worker as a proxy for its computer investment per worker.21 However, imports do not account for a country’s computer investments sourced from a domestic computer industry; this is an issue that becomes more salient in the later years of the dataset.22
A more optimal indicator would estimate computer access and usage among businesses, since such economic activity is more likely to produce productivity improvements than household use. I examined a few alternatives. The CHAT dataset employs the number of personal computers per capita, which is one of three measures highlighted by the authors as conveying information on GPTs.23 However, this indicator still does not capture the degree of computerization in productive processes, as opposed to personal use, and has limited temporal coverage compared to the ITU’s household computerization measure.24 The OECD collects some data on ICT access and usage by businesses, but this effort did not start until 2005 and covers only OECD countries.25
Fortunately, it stands to reason that a country’s household computer adoption can serve as a proxy for its computerization rates in business activities. In the appendix, I provide further support for this claim. Comparing the available data on household and business computerization for twenty-six countries between 2005 and 2014, I find a strong correlation between these two variables (correlation coefficient = 0.8).26
Summary of Main Model Specifications
To review, this chapter tests whether GPTs diffuse more intensively and quickly in countries that have institutional advantages in widening the pool of relevant engineering skills and knowledge. The first hypothesis reads as follows:
H1: Countries with higher levels of GPT skill infrastructure in software engineering will sustain more intensive computerization rates.
With country-years as the unit of analysis, I estimate time-series cross-sectional (TSCS) models of nineteen countries over twenty-six years. Quantitative analysis permits an expansion of scope beyond the great powers covered in the case studies. As outlined in the theory chapter, differences in GPT skill infrastructure are most relevant for economies that possess the absorptive capacity to assimilate new breakthroughs from the global technological frontier.27 Since less-developed economies are often still striving to build the baseline physical infrastructure and knowledge context to access the technological frontier, variation in GPT skill infrastructure among these countries is less salient. Thus, I limit the sample to nineteen G20 countries (the excluded member is the European Union), which represent most of the world’s major industrialized and emerging economies.
Before constructing TSCS regressions, I first probe the relationship between GPT skill infrastructure in software engineering and computerization rates. Prior to the inclusion of any control variables, I plot the independent and dependent variable in aggregate to gauge whether the hypothesized effect of GPT skill infrastructure is plausible. The resulting bivariate plot suggests that there could be a strong, positive relationship between these two variables (figure 6.2).28
The basic trend in figure 6.2 provides evidence for the contention that countries better equipped with computer science skill infrastructure experience higher rates of computerization. Although these preliminary results point to a strong relationship between these two variables, further tests are needed to rule out unseen confounders that could create spurious correlations and influence the strength of this relationship. TSCS regression analysis facilitates a deeper investigation of the relationship between GPT skill infrastructure and computerization.
To control for factors that could distort the relationship between computer-related skill infrastructure and computerization, I incorporate a number of control variables in the baseline model. Rich countries may be able to spend more on computer science education; additionally, they also more easily bear the expenses of adopting new technologies, as exemplified by large disparities between developed and developing countries in information technology investment levels.29 The inclusion of GDP per capita in the model accounts for economic development as a possible confounder. I use expenditure-side real GDP at current purchasing power parities (PPPs), which is best suited to comparing relative living standards across countries. A country’s total population, another control variable, addresses the possibility that larger countries may be more likely to benefit from network effects and economies of scale, which have been positively linked to technology adoption.30 I also include the polity score for regime type. Research suggests that democratic governments provide more favorable environments for technology diffusion, and studies have confirmed this connection in the specific context of internet technologies.31
Finally, the baseline model includes two control variables that represent alternative theories of how technological changes differentially advantage advanced economies. First, I include military spending as a proportion of GDP in the regressions. The case studies have interrogated the influential view that military procurement is an essential stimulus for GPT adoption.32 By examining the relationship between military spending and computerization across a large sample of countries, the statistical analysis provides another test of this argument. Moreover, the varieties of capitalism (VoC) scholarship suggests that liberal market economies (LMEs) are especially suited to form the general skills that could aid GPT adoption across sectors. Therefore, the baseline model also controls for whether a country is designated as an LME according to the VoC typology.33
In terms of model specification, I employ panel-corrected standard errors with a correction for autocorrelation, a typical method for analyzing TSCS data.34 Given the presence of both autocorrelation and heteroskedasticity, I estimate linear models on panel data structures using a two-step Prais-Winsten feasible generalized least squares procedure.35
Time-Series Cross-Sectional Results
Table 6.1 gives the results of the three initial models, which provide further support for the theoretical expectations.36 Model 1 incorporates controls that relate to economic size and level of development. Model 2 adds a control variable for regime type. Lastly, model 3 includes a variable that represents a prominent alternative theory for how technological breakthroughs can differentially advantage certain economies. This also functions as the baseline model. In all three models, the coefficient on the GPT skill infrastructure measure is positive and highly statistically significant (p < .05).
Table 6.1 Results of Time-Series Cross-Sectional Models
Dependent Variable
Computerization
(1)
(2)
(3)
GPT skill infrastructure
3.760**
4.064***
4.227**
(1.643)
(1.676)
(1.666)
GDP per capita
29.754***
29.319***
29.435***
(3.760)
(3.737)
(3.789)
Total population
6.969***
7.046***
6.781***
(1.625)
(1.654)
(1.549)
Polity score
−0.456
−0.472*
(0.295)
(0.277)
Military spending
−0.940
(2.413)
Liberal market economy
−2.194
(3.961)
Constant
−374.599***
−368.051***
−361.885***
(60.452)
(61.173)
(58.374)
Observations
383
370
370
Note: Standard errors in parentheses.
*p < .10; **p < .05; ***p < .01
The effect of GPT skill infrastructure on GPT adoption is also substantively significant. Given the coefficient of the GPT skill infrastructure measure in the baseline model,37 a 1 percent increase in the universities per 100,000 people that provide software engineering education results in an increase of the computerization rate by 0.042 percentage points.38 Though the substantive effect seems small at first glance, its magnitude becomes clear when contextualized by differences in GPT skill infrastructure across the sample. For example, in China over this time period, the average number of universities per 100,000 people that met my baseline for GPT skill infrastructure was 0.040. The corresponding figure for the United States was 0.248. According to the coefficient estimate for GPT skill infrastructure, this difference of 520 percent corresponds to a difference of nearly 22 percentage point units in the computerization rate.
It should be noted that only two control variables, economic development and population, came in as statistically significant in the baseline model. As expected, wealthier countries and more populous countries presided over more intensive adoption of computing technologies. The null result for regime type is worth highlighting, as the effects of democracy on technology adoption are disputed.39 Finally, contrary to the expectations of competing explanations to GPT diffusion theory, the effects of military spending and VoC are insignificant. This is consistent with the findings from the historical case studies.
Quantitative appendix table 1 displays the results after incorporation of three additional controls. First, trade linkages expose countries to advanced techniques and new ideas, opening the door to technology diffusion. A high level of trade openness has been associated with more intensive adoption of information technologies.40 Relatedly, there is evidence that a country’s openness to international trade has a positive and sizable effect on various measures of innovation, including high-technology exports, scientific publications, and patents.41 Second, higher urban density has been linked to faster diffusion of technologies such as the television and the internet.42 Model 8 incorporates a trade openness variable and an urbanization variable.
Third, patterns at the regional level could shape how computerization spreads around the world. Scholars have identified such regional effects on the diffusion of ideas, policies, and technologies.43 In model 9, I assess spatial dynamics with dummy variables for the following regions: East Asia and Pacific; Europe and Central Asia; Latin America and Caribbean; the Middle East and North Africa; North America; South Asia; and sub-Saharan Africa.44 The positive effect of GPT skill infrastructure on computerization stays strong and highly statistically significant across these two models.
To ensure that the results were not determined by my choice of independent variable, I constructed an alternative specification of GPT skill infrastructure. Reanalyzing data on 467,198 software engineering publications, I counted the number of distinct authors for each country, as a proxy for the breadth of researchers who could foster human capital in software engineering. Though I still maintain that the primary specification best captures software engineering skill infrastructure, this alternative construction guards against possible issues with institution-based indicators, such as problems with institutional disambiguation and nonstandardized author affiliations.45 With this alternative specification, the estimated effect of GPT skill infrastructure on computerization remains positive and significant for the baseline model as well as for models with additional controls.46
Duration Analysis
When it comes to whether great powers can harness the potential of GPTs for productivity growth, the speed of adoption—not just the intensity of adoption—is pertinent. In the historical case studies, technological leaders adapted more quickly to industrial revolutions because of their investments in widening the base of engineering knowledge and skills associated with GPTs. This leads to the second hypothesis.
H2: Countries with higher levels of GPT skill infrastructure in software engineering will experience faster levels of computer adoption.
In testing this hypothesis, the dependent variable shifts to the amount of time it takes for a country to reach a particular computerization rate. A critical step is to establish both the specific computerization rate that constitutes successful “adoption” of computers as well as when the process of diffusion begins. Regarding the former, I count the “first adoption” of computerization as when the proportion of households with a computer in a country reaches 25 percent. This approach is in line with Everett Rogers’s seminal work on the S-shaped curve for successful diffusion of an innovation, which typically takes off once the innovation reaches a 10 to 25 percent adoption rate.47 For the duration analysis, since many of the countries enter the dataset with levels of computer adoption higher than 10 percent, the 25 percent level threshold is more suitable.48
I take 1995 as the starting point for the diffusion of computers as a GPT. Though an earlier date may be more historically precise, the 1995 date is more appropriate for modeling purposes, as the data on computerization rates for countries before this time are sparse. In a few cases, a country clearly achieved the 25 percent computerization threshold before 1995.49 As a practical measure to estimate the duration models, I assume that the time it took for these countries to adopt computers was one year. Right-censoring occurs with the last year of data, 2020, as many countries still had not reached the 25 percent computerization rate.
Using these data, I employ a Cox proportional hazards model to estimate the time it takes for countries to reach a 25 percent computerization rate based on the start date of 1995. Often used by political scientists to study conflict duration or the survival of peace agreements, duration models are also commonly used to investigate the diffusion of new technologies and to determine why some firms take longer to adopt a certain technology than others.50 Freed from the demands of TSCS analysis for yearly data, the duration analysis expands the county coverage, incorporating all upper-middle-income economies or high-income economies, based on the World Bank’s income group classifications.51 The resulting sample, which excludes countries that never attained the 25 percent computerization threshold, includes seventy-six countries.
Table 6.2 reports the estimated coefficients from the duration analysis. Positive coefficients match with a greater likelihood of reaching the computerization threshold. I use the same explanatory variable and controls as the baseline model from the TSCS analysis. These variables all enter the model with their measures in 1995. Model 4a takes the 25 percent computerization rate as the adoption threshold, while model 4b adjusts it to 20 percent to ensure that this designation is not driving results.
Table 6.2 Time to Computerization by Country
Dependent Variable
25% Threshold(4a)
20% Threshold(4b)
GPT skill infrastructure
0.673***
0.517***
(0.137)
(0.119)
GDP per capita
1.186***
1.110***
(0.335)
(0.288)
Total population
0.127
0.062
(0.085)
(0.074)
Polity score
0.022
0.024
(0.025)
(0.023)
Military spending
0.017
−0.042
(0.218)
(0.198)
Liberal market economy
0.760
0.785
(0.503)
(0.493)
N (number of events)
76 (74)
83 (83)
Likelihood ratio test (df = 6)
112.9***
111.2***
Note: Standard errors in parentheses.
*p < .10; **p < .05; ***p < .01
As the models demonstrate, the effect of GPT skill infrastructure on the speed by which countries achieve computerization is positive and highly statistically significant, providing support for hypothesis 2. Based on model 4a’s hazard ratio for the independent variable (1.96) for a given year, a tenfold increase in software engineering university density doubles the chances of a country reaching the computerization threshold.52 These results hold up after introducing additional control variables (quantitative appendix table 3).53
A Cross-Sectional Approach: Averages across the 1995–2020 Period
As an additional check, I collapse the panel dataset into cross-sectional averages of GPT skill infrastructure and computerization over the 1995–2020 period in a large sample of countries. In certain aspects, cross-sectional evidence could be more appropriate for comprehending the impact of features, like skill formation institutions, that are difficult to capture in yearly intervals because they change gradually.54 This approach allows for more countries to be included, as the yearly data necessary for TSCS analysis were unavailable for many countries. Limiting the sample based on the same scope conditions as the duration analysis leaves 127 countries.
I also include the same set of controls used in the previous analyses of the panel data: GDP per capita, total population, regime type, military spending, and liberal market economies. I employ ordinary least squares (OLS) regression to estimate the model. Since both a scale-location plot and a Breausch-Pagan test demonstrate that heteroskedasticity is not present in the data, it is appropriate to use an OLS regression estimator with normal standard errors.
The results of the regression analysis provide further support for the theoretical expectations. To recap, the independent variable is the estimated average skill infrastructure for software engineering between 1995 and 2020, and the dependent variable is the average computerization rate during the same period. Since analyzing bibliographic information on yearly software engineering publications for 127 countries is a demanding exercise, I estimated the average number of universities that nurture software engineering skills based on publications for the two middle years (2007, 2008) in the dataset, instead of deriving this average based on publication data across the entire time range.55 Table 6.3 displays the results, with the incremental inclusion of control variables, also averaged over the period 1995–2020, across three models.56 The coefficient on the GPT skill infrastructure measure remains positive and highly statistically significant across all three models (p < .01).
Table 6.3 GPT Skill Infrastructure Predicts More Computerization
Dependent Variable
Computerization
(5)
(6)
(7)
GPT skill infrastructure
3.211***
3.737***
3.761***
(0.528)
(0.609)
(0.649)
GDP per capita
16.617***
15.536***
14.977***
(1.564)
(1.723)
(1.812)
Total population
−1.647***
−0.739
−0.831*
(0.440)
(0.485)
(0.500)
Polity score
−0.066
−0.070
(0.147)
(0.180)
Military spending
0.577
(1.604)
Liberal market economy
5.017
(4.269)
Constant
−83.099***
−86.165***
−79.773***
(20.577)
(22.316)
(23.636)
Observations
127
112
110
R2
0.812
0.833
0.834
Note: Standard errors in parentheses.
*p < .10; **p < .05; ***p < .01
I perform several additional tests to confirm the robustness of the results. I first include the same additional controls used in the preceding TSCS analysis. Quantitative appendix table 4 shows that the main findings are supported. One limitation of models that rely on cross-sectional averages is endogeneity arising from reverse causality. In other words, if greater diffusion of computers throughout the economy spurs more investment in institutions that broaden the pool of software engineers, then this could confound the baseline model’s estimates. To account for this possibility, in model 16 in quantitative appendix table 5, I operationalize GPT skill infrastructure using the estimate for the year 1995, the start of the period, instead of its average level over the 1995–2020 period.57 Thus, this model captures the impact of GPT skill infrastructure in 1995 on how computerization progressed over the remaining sample years.58 The effect remains positive and statistically significant.
While this chapter’s primary aim is to investigate empirical patterns expected by GPT diffusion theory, the quantitative analysis can also probe whether computerization is positively influenced by institutions that complement LS product cycles. To that end, I add a control variable that stands in for the institutional competencies prioritized by the LS model.59 Measures of computer exports and ICT patents serve as two ways to capture a country’s ability to generate novel innovations in the computer industry.60 In the resulting analysis, the LS-linked variables do not register as statistically significant.61
These results should be interpreted with care. In many cases, measures of institutional capacity to build a strong, innovative computer sector may be highly correlated with measures of GPT skill infrastructure. Because the statistical approach struggles to differentiate between the causal processes that connect these two features and computerization, the historical case studies take on the main burden of comparing the GPT diffusion and LS mechanisms against each other. Still, the inclusion of variables linked with the LS mechanism does suggest that, in the context of computerization, there is ineffectual evidence that the presence of a strong leading sector spills over into other sectors and generates multiplier effects—a key observable implication of LS accounts.62 Additionally, these models drive home the importance of differentiating between institutions linked to innovative activities (in the sense of introducing new products and processes) and engineering-oriented institutions, like GPT skill infrastructure, which are more connected to adoptive activities.63
Summary
Using a variety of statistical methods, this chapter tested the expectation that countries better equipped to widen the base of engineering talent in a GPT will be more successful at diffusing that GPT throughout their economies. The combination of TSCS models, duration analysis, and cross-sectional regressions lends credence to the strength of the relationship between GPT skill infrastructure and computerization. The results hold across a range of additional tests and robustness checks.
There are two major limitations to this chapter’s approach. First, the statistical analysis should be interpreted mainly as an independent evaluation of GPT diffusion theory, not as an additional comparison between GPT diffusion theory and the causal pathway linked to LS product cycles. In the large-scale statistical analysis, indicators linked with the LS mechanism can also be associated with higher computerization rates, making it difficult to weigh the two explanations against each other. The rich historical detail in the case studies therefore provides the prime ground for tracing causal mechanisms.
Second, this chapter evaluates only one aspect of GPT skill infrastructure. A more comprehensive assessment would include not just the capacity to widen the pool of software engineering talent, which was the independent variable in this analysis, but also the strength of information flows between the GPT sector and application sectors. For instance, in the IR-2 case, both the United States and Germany trained large numbers of mechanical engineers, but American technological institutes placed more emphasis on practical training and shop experience, which strengthened connections between the US mechanical engineering education system and industrial application sectors. Building on data collection efforts that are starting to measure these types of linkages, such as the proportion of publications in a technological domain that involve industry-academia collaborations, future research should conduct a more complete assessment of GPT skill infrastructure.64
Notwithstanding these limitations, the quantitative analysis backs a key observable implication of GPT diffusion theory: advanced economies’ level of GPT skill infrastructure is strongly linked to GPT adoption rates. Not only does this provide some initial support for the generalizability of this book’s central argument beyond just great powers, but it also gives further credibility to GPT diffusion theory’s relevance to US-China competition today.
7 US-China Competition in AI and the Fourth Industrial Revolution
THE FIRST MACHINE to defeat a human Go champion; powerful language models that can understand and generate humanlike text; a computer program that can predict protein structures and enable faster drug discovery—these are just a few of the newest discoveries in AI that have led some to declare the arrival of a Fourth Industrial Revolution (IR-4).1 As for the latest geopolitical trends, China’s rise has been the dominant story of this century as national security establishments grapple with the return of great power competition. Located squarely at the intersection of these two currents, the US-China technological rivalry has become an inescapable topic of debate among those interested in the future of power—and the future itself.
Who will lead the way in the Fourth Industrial Revolution? To answer this question, leading thinkers and policymakers in the United States and China are both drawing lessons from past technology-driven power transitions to grapple with the present landscape. Unfortunately, as this chapter argues, they have learned the wrong lessons. Specifically, the leading-sector perspective holds undue influence over thinking about the relationship between technological change and the possibility of a US-China power transition. Yet careful tracing of historical cases and statistical analysis have revealed that the GPT mechanism provides a better model for how industrial revolutions generate the potential for a power transition. When applied to the IR-4 and the evolving US-China power relationship, GPT diffusion theory produces different insights into the effects of the technological breakthroughs of today on the US-China power balance, as well as on the optimal strategies for the United States and China to pursue.
This chapter sketches out the potential impacts of today’s emerging technologies on the US-China power balance. I first describe the current productivity gap between the United States and China, with particular attention to concerns that the size of this gap invalidates analogies to previous rising powers. Next, I review the array of technologies that have drawn consideration as the next GPT or next LS. Acknowledging the speculative nature of technological forecasting, I narrow my focus to developments in AI because of its potential to revitalize growth in ICT industries and transform the development trajectories of other enabling technologies.
The essence of this chapter is a comparison of the implications of the LS and GPT mechanisms for how advances in AI will affect a possible US-China economic power transition. In contrast to prevailing opinion, which hews closely to the LS template, GPT diffusion theory suggests that the effects of AI on China’s rise will materialize through the widespread adoption of AI across many sectors in a decades-long process. The institutional factors most pertinent to whether the United States or China will more successfully benefit from AI advances are related to widening the base of AI-related engineering skills and knowledge. I also spell out how the implications of GPT diffusion theory for the US-China power balance differ from those derived from alternative explanations.
The objective here is not to debate whether China will or will not catch up to the United States, or whether technological capabilities are more significant than all other considerations that could affect China’s long-term economic growth. Rather, this chapter probes a more limited set of questions: If emerging technologies were to significantly influence the US-China economic power balance, how would this occur? Which country is better positioned to take advantage of the Fourth Industrial Revolution? What would be the key institutional adaptations to track?2
A Power Transition in Progress?
Over the past four decades, there has been no greater shift in the global economic balance than China’s rise. China is either already the world’s largest economy, if measured by purchasing power parity (PPP) exchange rates, or is projected to soon overtake the United States, based on nominal exchange rates.3 China’s impressive economic growth has led many to proclaim that the era of US hegemony is over.4
Economic size puts China in contention with the United States as a great power competitor, but China’s economic efficiency will determine whether a power transition occurs. Countries like Switzerland outpace the United States on some measures of economic efficiency, but they lack the economic size to contend. Other rising powers, such as India, boast large economies but lag far behind in economic efficiency. Mike Beckley concludes: “If the United States faces a peer competitor in the twenty-first century … it will surely be China.”5 For this conditional to be true, China’s productivity growth is critical.6 This is not the first time in history that China has boasted the world’s largest economy; after all, it held that distinction even as Britain was taking over economic leadership on the back of the First Industrial Revolution.
Where does China currently stand in comparison to the productivity frontier? Based on 2018 figures, China’s real GDP per capita (at 2010 PPPs) is about 30 percent that of the United States.7 From 2000 to 2017, China’s total factor productivity (TFP) never surpassed 43 percent of US TFP (figure 7.1).8 In 2015, labor productivity in China remained at only 30 percent of that in the United States, though this figure had doubled over the past two decades.9
These numbers suggest that China sits much further from the productivity frontier than past rising powers. If the US-China power relationship is fundamentally different from those in previous eras, the relevance of conclusions from previous cases could be limited.10 For instance, in the early years of the IR-1, Britain was only slightly behind the Netherlands, the productivity leader at the time. The United Kingdom’s GDP per capita was 80 percent of Dutch GDP per capita in 1800.11 Similarly, at the beginning of the IR-2, the United States trailed Britain in productivity by a small margin. In 1870, labor productivity and TFP in the United States were 90 percent and 95 percent, respectively, of labor productivity and TFP in Britain.12 During the 1870s, average GDP per capita in the United Kingdom was about 15 percent higher than average GDP per capita in the States.13
FIGURE 7.1. US-China Productivity Gap (2000–2017). Source: Penn World Table version 9.1; Feenstra, Inklaar, and Timmer 2015.
Still, that China could surpass the United States in productivity leadership is not outside the realm of possibility.14 The IR-3 case is a better comparison point for the current productivity gap between the United States and China. In 1960, the start of the IR-3 case, Japanese GDP per capita was 35 percent of US GDP.15 At the time, Japan’s TFP was 63 percent of US TFP, and Japan’s labor productivity was only 23 percent of US labor productivity, a lower proportion than the China-US ratio at present.16 Despite the initial chasm, the TFP gap between the United States and Japan narrowed to only 5 percent in 1991.17
Indeed, productivity growth is crucial for China to sustain its economic rise in the long term. For the 1978–2007 period, Xiaodong Zhu decomposed the sources of China’s economic growth into labor deepening, human capital, capital deepening, and total factor productivity growth. He found that growth in TFP accounted for 78 percent of China’s growth in GDP per capita.18 The burden on TFP improvements will only increase, given the diminishing impact of other drivers of China’s growth miracle, such as urbanization and demographic dividends.19
Whether China can sustain productivity growth is an open question. Plagued by inefficient infrastructure outlays, China’s aggregate TFP growth declined from 2.8 percent in the decade before the global financial crisis to 0.7 percent in the decade after (2009–2018).20 If calculated using alternative estimates of GDP growth, China’s TFP growth was actually negative from 2010 to 2017, averaging −0.5 percent.21 This productivity slowdown is not unique to China. Even before the 2008 global financial crisis, there was a slowdown in TFP growth among advanced economies due to waning effects from the information and communications technologies (ICTs) boom.22 China’s labor productivity growth also decelerated from 8.1 percent in 2000–2007 to 4.2 percent in 2011–2019, but the later period’s slowed growth rate was still six times greater than the US labor productivity growth rate in the same period.23
Adaptation to technological advances will be central to China’s prospects of maintaining high rates of productivity growth. China’s leaders worry about getting stuck in the “middle-income trap,” a situation in which an economy is unable to advance to high-income status after it exhausts export-driven, low-cost manufacturing advantages. Many studies have stressed the linkage between China’s capacity to develop and absorb emerging technologies and its prospects for escaping the middle-income trap.24 The Chinese government also increasingly pushes the development and adoption of information technology and other cutting-edge technologies as a way to increase TFP.25 Thus, tracking China’s future productivity growth necessitates a better understanding of specific technological trajectories in the current period.
Key Technological Changes in the IR-4
ICTs, the key technological drivers of the IR-3, are still at the heart of the IR-4. From visionaries and daydreamers to economists and technology forecasters, there is a wide-ranging consensus that AI will breathe new life into the spread of digitization. The World Economic Forum calls AI the “engine that drives the Fourth Industrial Revolution.”26 Kai-Fu Lee, the former head of Google China, boldly asserts, “The AI revolution will be on the scale of the Industrial Revolution but probably larger and definitely faster.”27 To further explore AI’s role in the IR-4, I examine this technological domain as a source of both GPT and LS trajectories.
Candidate Leading Sectors
LS accounts forecast that in future waves of technological change ICTs will continue to drive economic transformation. According to one analysis of US-China technological rivalry in the twenty-first century, ICTs are “widely regarded as the current leading sector.”28 I reviewed five key texts that predicted future leading sectors, all written by scholars who study historical cycles of technological change and global leadership transitions.29 These forecasts also highlight other candidate leading sectors, including lasers and new sources of energy, but they converge on ICTs as the leading sector of the next wave of technological disruption.
Informed by the LS model, AI’s effects on global technological competition are often framed through its potential to open up new opportunities for latecomers to catch up and leapfrog advanced countries in key segments like AI chips. China’s national AI development plan outlines its ambition to become the world’s leading center of innovation in AI by 2030.30 Scholars analyze China’s capacity to develop global intellectual monopolies in certain AI applications and enhance independent innovation in AI so as to guard against other countries leveraging weaponized interdependence.31 Descriptions of China’s AI strategy as aimed toward seizing “the commanding heights” of next-generation technologies reflect the belief that competition in AI will be over global market shares in strategic sectors.32
Candidate GPTs
Among the possible GPTs that could significantly impact a US-China economic power transition, AI stands out. Like the literature on leading sectors, the GPT literature also converges on ICTs as a continued driver of technological revolution. Kenneth Carlaw, Richard Lipsey, and Ryan Webb, three pioneers of GPT-based analysis, identify programmable computing networks as the basic GPT that is driving the modern ICT revolution.33 Crucially, AI could open up a new trajectory for this ICT revolution. Recent breakthroughs in deep learning have improved the ability of machines to learn from data in fundamental ways that can apply across hundreds of domains, including medicine, transportation, and other candidate GPTs like biotechnology and robotics. This is why AI is often called the “new electricity”—a comparison to the prototypical GPT. Economists regard it as the “next GPT”34 and “the most important general-purpose technology of our era.”35
Several studies have found evidence for a GPT trajectory in AI. One study, using a novel dataset of preprint papers, finds that articles on deep learning conform with a GPT trajectory.36 Using patent data from 2005 to 2010 to construct a three-dimensional indicator for the GPT-ness of a technology, Petralia ranks technological classes based on their GPT potential.37 His analysis finds that image analysis, a field that is closely tied to recent advances in deep learning and AI, ranks among the top technological classes in terms of GPT-ness.38 Another effort employs online job posting data to differentiate among the GPT-ness of various technological domains, finding that machine learning technologies are more likely to be GPTs than other technologies such as blockchain, nanotechnology, and 3D printing.39
To be sure, forecasts of future GPTs call attention to other technological trends as well. Other studies have verified the GPT potential of biotechnology.40 Robotics, another candidate GPT for the IR-4, could underpin “the next production system” that will boost economy-wide productivity, succeeding the previous one driven by information technology.41 While my primary reasons for limiting my analysis to AI are based on space constraints as well as on its prominence in the surrounding literature, it is also important to note that developments in both biotechnology and robotics are becoming increasingly dependent on advances in deep learning and big data.42
The Limits of Technological Foresight
Unlike exercises to pinpoint key technologies of previous industrial revolutions, which benefited from hindsight, identifying technological drivers in the IR-4 is a more speculative exercise. It is difficult to find true promise amid the hype. The task is made harder by the fact that even experts and technological forecasting bodies regularly miss the next big thing. In 1945, a team led by Dr. Theodore von Kármán, an eminent aerospace engineer, published Toward New Horizons, a thirty-two-volume text about the future of aviation. The study failed to foresee major new horizons such as the first human in space, intercontinental ballistic missiles (ICBMs), and solid-state electronics—all of which emerged within fifteen years.43 In the early 1990s, the US Army conducted a technology forecast assessment to identify the technologies most likely to transform ground warfare in the next century. When the forecast was evaluated in 2008 by the Army’s senior scientists and engineers, it graded out at a “C.” Among its most significant misses was the development of the internet.44
I am no Cassandra. It is very possible that if I were writing this book in 2000, this chapter would focus on the promise of nanotechnology, not AI. At that time, President Bill Clinton had just unveiled the National Nanotechnology Initiative. In a 2003 speech, Philip J. Bond, the undersecretary for technology at the Department of Commerce at the time, declared:
Nano’s potential rises to near Biblical proportions. It is not inconceivable that these technologies could eventually achieve the truly miraculous: enabling the blind to see, the lame to walk, and the deaf to hear; curing AIDS, cancer, diabetes and other afflictions; ending hunger; and even supplementing the power of our minds, enabling us to think great thoughts, create new knowledge, and gain new insights.45
Decades later, there is a collective exhaustion around the hype surrounding nanotechnology, a phenomenon one scientist calls “nanofatigue.”46
One lesson that stands out from mapping the technological landscape in past industrial revolutions is that the most significant GPTs of an era often have humble origins. In the IR-2, new innovations in electricity and chemicals garnered the most attention, but America’s economic rise owed more to advances in machine tools that were first introduced decades earlier. In the same way, “old” GPTs like electricity could still shock the world.47 Today there is still a lot of potential for expanded industrial electrification, which could have a major impact on productivity.48 Similarly, high-capacity battery technologies could transform productivity on a broad scale.49 Interestingly, patent data also demonstrate the continued importance of electrical technologies. Among the top ten GPT candidates as ranked by Petralia’s indicator of “GPT-ness,” there were as many technological classes in the electrical and electronic category as there were in the computers and communications category.50
For my purposes, it is reassuring that developments in AI also draw on a long history. In the United States, the legitimization of AI as an important field of research dates back to the 1960s.51 Thus, though the rest of the chapter takes AI as the most important GPT for the near future, it does so with a humble mindset, acknowledging that looking forward to the future often starts with digging deeper into the past.
The GPT vs. LS Mechanisms in the IR-4
There has been no shortage of speculation about whether the United States or China is better fit for the new AI revolution. Each week it seems there is a new development in the “AI arms race” between the two nations.52 Many believe that China is an AI superpower on the verge of overtaking the United States in the key driver of the IR-4.53 As the following sections will show, these discussions tend to follow the LS template in their assumptions about the trajectory of AI development and key institutional adjustments.
Conversely, GPT diffusion theory provides an alternative model for how AI could affect the US-China power balance, with implications for the optimal institutional adaptations to the AI revolution. I conclude that, if the lessons of past industrial revolutions hold, the key driver of a possible US-China economic power transition will be the relative success of these nations in diffusing AI throughout their economies over many decades. This technological pathway demands institutional adaptations to widen the base of AI engineering skills and knowledge. While recognizing that international competition over AI is still in the early innings, this chapter outlines a preliminary framework for assessing which country’s roster is better equipped for success.
Impact Timeframe: The Decisive Years in the US-China AI Competition
If guided by the LS mechanism, one would expect the impacts of AI on US-China power competition to be very significant in the early stages of the technology’s trajectory. Indeed, many prominent voices have articulated this perspective. Consider, for example, a report titled “Is China Beating the US to AI Supremacy?,” authored by Professor Graham Allison, the director of Harvard Kennedy School’s Belfer Center for Science and International Affairs and Eric Schmidt, former CEO of Google and cochair of the National Security Commission on Artificial Intelligence (NSCAI). For Allison and Schmidt, the decisive years in US-China AI competition are just around the corner. Assuming that AI advances will be rapidly adopted across many economic domains, their aim is to “sound an alarm over China’s rapid progress and the current prospect of it overtaking the United States in applying AI in the decade ahead.”54 Shaped by a similar framework, other influential texts also predict that China’s productivity boost from AI will come to fruition in the 2020s.55
If GPT diffusion theory serves as the basis for analysis, these influential texts severely underestimate the time needed for economic payoffs from AI. Historical patterns of GPT advance have borne out that, even in early adopter countries, it takes at least three or four decades for these fundamental technologies to produce a significant productivity boost.56
Using this pattern as guidance, we can roughly project AI’s impact timeframe, after establishing an initial emergence date for this GPT. In 2012, the AlexNet submission to ImageNet, a competition that evaluates algorithms on large-scale image classification, is widely recognized as spurring this current, deep learning–based paradigm of AI development.57 If using the metric of when a GPT achieves a 1 percent adoption rate in the median sector, the AI era probably began in the late 2010s.58 As of 2018, according to the most recent census survey on the extent of AI adoption in the US economy, only 2.75 percent of firms in the median sector reported using AI technologies.59 Thus, regardless of which arrival date is used, if AI, like previous GPTs, requires a prolonged period of gestation, substantial productivity payoffs should not materialize until the 2040s and 2050s.60
Of course, other factors could affect AI’s expected impact timeframe, including the possibility that the general process of technological adoption is accelerating. Some evidence indicates that the waiting time for a significant productivity boost from a new GPT has decreased over time.61 Lee argues that the AI revolution will be faster than previous GPT trajectories owing to the increasingly frictionless distribution of digital algorithms and more mature venture-capital industry.62 Nevertheless, preliminary evidence suggests that AI will face similar implementation lags as previous GPTs, including bottlenecks in access to computing resources, human capital training, and business process transformations.63
Phase of Relative Advantage: Innovation-centrism and China’s AI Capabilities
Debates about China’s scientific and technological power reduce complex dynamics to one magic word—“innovation.”64 Whether China can generate novel technologies is often the crux of debates over China’s growing scientific and technological capabilities and a potential US-China power transition.65 For David Rapkin and William Thompson, the prospect of China overtaking the United States as the leading power is dependent on “China’s capacity to innovate”—specifically as it relates to revolutionary technological changes that allow challengers to leapfrog the leader in economic competition.66 “If … China’s innovativeness continues to lag a considerable distance behind that of the US, then China overtaking the US might wait until the twenty-second century,” they posit.67 China’s innovation imperative, as Andrew Kennedy and Darren Lim describe it in language common to LS analysis, is motivated by “monopoly rents generated by new discoveries.”68
Innovation-centric views of China’s AI capabilities paint an overly optimistic picture of China’s challenge to US technological leadership. Allison and Schmidt’s Belfer Center paper, for instance, emphasizes China’s growing strength in AI-related R&D investments, leading AI start-ups, and valuable internet companies.69 Likewise, the NSCAI’s final report suggests that China is poised to overtake the United States in the capacity to generate new-to-the-world advances in AI, citing shares of top-cited, breakthrough papers in AI and investments in start-ups.70 These evaluations match up with viewpoints that are bullish on China’s overall technological capabilities, which also point to its impressive performance along indicators of innovation capacity, such as R&D expenditures, scientific publications, and patents.71
Some other comparisons of US and Chinese AI capabilities arrive at the opposite conclusion but still rely on the LS template. For instance, two Oxford scholars, Carl Frey and Michael Osborne, have likened claims that China is on the verge of overtaking the United States in AI to overestimates of Japan’s technological leadership in computers in the 1980s. In their view, just like Japan, China will fail to overtake the United States as the world’s technological leader because of its inability to produce radical innovations in AI. In fact, they claim, the prospects are even bleaker this time around: “China, if anything, looks less likely to overtake the United States in artificial intelligence than Japan looked to dominate in computers in the 1980s.”72
If analysis of US-China competition in AI was centered on GPT diffusion theory, it would focus more on China’s capacity to widely adopt AI advances. In this scenario, it is neither surprising nor particularly alarming that China, like other great power contenders such as Japan in the IR-3, Germany in the IR-2, and France in the IR-1, contributes to fundamental innovations. No one country will corner all breakthroughs in a GPT like AI. The key point of differentiation will be the ability to adapt and spread AI innovations across a wide array of sectors.
A diffusion-centric perspective suggests that China is far from being an AI superpower. Trends in ICT adoption reveal a large gap between the United States and China. China ranks eighty-third in the world on the International Telecommunication Union’s ICT development index, a composite measure of a country’s level of networked infrastructure, access to ICTs, and adoption of ICTs.73 By comparison, the United States sits among the world’s leaders at fifteenth. Though China has demonstrated a strong diffusion capacity in consumer-facing ICT applications, such as mobile payments and food delivery, Chinese businesses have been slow to embrace digital transformation.74
In fact, it is often Chinese scholars and think tanks that acknowledge these deficiencies. According to an Alibaba Research Institute report, China significantly trails the United States in penetration rates of many digital technologies across industrial applications, including digital factories, industrial robots, smart sensors, key industrial software, and cloud computing.75 China also significantly trails the United States in an influential index for adoption of cloud computing, which is essential to implementing AI applications.76 In 2018, US firms averaged a cloud adoption rate of over 85 percent, more than double the comparable rate for Chinese firms.77
To be fair, China has achieved some success in adopting robots, a key application sector of AI. China leads the world in total installations of industrial robots. Aided by favorable industry composition and demographic conditions, China added 154,000 industrial robots in 2018, which was more than were installed by the United States and Japan combined.78 Based on 2021 data from the International Federation of Robotics, China outpaces the United States in robot density as measured by the number of industrial robots per 10,000 manufacturing employees.79
However, China’s reputed success in robot adoption warrants further scrutiny. The IFR’s figures for employees in China’s manufacturing sector significantly underestimate China’s actual manufacturing workforce. If these figures are revised to be more in line with those from the International Labor Organization (ILO), China’s robot density would fall to less than 100 robots per 10,000 manufacturing employees, which would be around one-third of the US figure.80 On top of that, talent bottlenecks hamper robot diffusion in China, since skilled technicians are required to reprogram robots for specific applications.81 An unused or ineffective robot counts toward robot density statistics but not toward productivity growth.
Breadth of Growth: Picking Winners vs. Horizontal Approaches to AI Development
Divergent perspectives on the breadth of growth in technological revolutions also separate LS-based and GPT-based views of the US-China case. If technological competition in the IR-4 is limited to which country gets a bigger share of the market in new leading industries like AI, then direct sectoral interventions in the mold of China’s AI strategy could be successful. However, if the breadth of growth in the IR-4 follows the GPT trajectory of the three previous industrial revolutions, another approach will be more effective.
China’s AI strategy has hewed closely to the LS model. This approach builds off a series of directives that prioritize indigenous innovation in select frontier technologies, an emphasis that first appeared in the 2006 “National Medium- and Long-Term Plan for the Development of Science and Technology” and extends through the controversial “Made in China 2025” plan.82 Since the mid-2000s, the number of sectoral industrial policies issued by the State Council, China’s cabinet-level body, has significantly increased.83 Appropriately, the State Council’s 2017 AI development plan outlined China’s ambitions to become the world’s primary innovation center for AI technology.84
On the breadth of growth dimension, tension between GPT diffusion theory and China’s application of the LS template is rooted in differing expectations for how the economic boost from revolutionary technologies will unfold. Take, for example, China’s 2010 “Strategic Emerging Industries” (SEI) initiative, which targets seven technological sectors based on opportunities for China to leapfrog ahead in new industries.85 Oriented around assumptions that a limited number of technologically progressive industries will drive China’s future growth, the SEI defines success based on the resultant size of these industries, as measured by their value added as a share of GDP.86
In contrast, GPT diffusion theory expects that, in the country that best capitalizes on the IR-4, productivity growth will be more dispersed. In this view, the AI industry never needs to be one of the largest, provided that AI techniques trigger complementary innovations across a broad range of industries. Relatedly, some Chinese thinkers have pushed back against industrial policies that favor narrow technology sectors. A research center under China’s own State Council, in a joint analysis with the World Bank, concluded in 2012: “A better innovation policy in China will begin with a redefinition of government’s role in the national innovation system, shifting away from targeted attempts at developing specific new technologies and moving toward institutional development and an enabling environment that supports economy-wide innovation efforts within a competitive market system.”87 The economy-wide transformation enabled by AI, if it lives up to its potential as a GPT, demands a more broad-based response.
When it comes to technology policy, there is always a push and pull between two ends of a spectrum. Vertical industrial policy, or “picking winners,” targets certain technologies, often leading to top-down intervention to ensure that the nation’s firms are competitive in specific industries. Horizontal industrial policy promotes across-the-board technological development and avoids labeling certain technologies as more significant than others. This book argues that both camps have it partly right, at least when it comes to ensuring long-term economic growth in times of technological revolution. Picking technological winners is needed in that some technologies do matter more than others; however, the “winners” are GPTs, which require horizontal industrial policies to diffuse across many application sectors. Institutions for skill formation in AI engineering, the subject of the next section, split the difference between these two approaches.
Institutional Complementarities: GPT Skill Infrastructure in the IR-4
In 2014, Baidu, one of China’s leading tech giants, hired Andrew Ng away from Google, poaching the cofounder of Google’s deep learning team. Three years later, Baidu lured Qi Lu away from Microsoft, where he had served as the architect of the company’s AI strategy. Their departures were headline news and spurred broader discussions about China’s growing AI talent.88
When Alibaba, another one of China’s tech giants, celebrated its listing on the Hong Kong stock exchange in November 2019, it showcased a different form of AI talent. In one picture of the gong-ringing celebration, Yuan Wenkai, who works for an Alibaba-owned logistics warehouse, stood third from the right. A former tally clerk who graduated from a run-of-the-mill Guangdong vocational school, Yuan holds substantial expertise in automation management. His success with boosting the sorting capacity of a logistics warehouse by twenty thousand orders per hour—responding to elevated demand during the shopping frenzy of Single’s Day (November 11)—merited an invite to the ceremony.89
Even as AI systems exceed human-level performance at tasks ranging from playing Go to translating news articles, human talent will remain crucial for designing and implementing such systems.90 According to one global survey of more than three thousand business executives, landing the “right AI talent” ranked as the top barrier to AI adoption for companies at the frontier of incorporating AI into their products, services, and internal processes.91 But what makes up the “right AI talent”? In its distilled form, GPT diffusion theory suggests that China’s chance of leading the AI revolution rests more on the Yuan Wenkais of the world than the Andrew Ngs. The most important institutional adjustments to the IR-4 are those that widen the pool of AI engineering skills and knowledge.
Indeed, alongside the maturation of the AI field, recent reports have emphasized skills linked to implementing theoretical algorithms in practice and in ways suited for large-scale deployment. In early 2022, the China Academy of Information and Communications Technology (CAICT), an influential research institute under the Ministry of Industry and Information Technology, published two reports that identified AI’s “engineering-ization” (工程化) as a significant trend that involves addressing challenges in transforming AI-based projects from prototypes to large-scale production.92 Relatedly, per Burning Glass job postings from 2010 to 2019, the overall increase in demand for “AI-adjacent” positions in the United States far exceeded that for “core AI” positions.93 Covering skills needed to implement AI throughout many sectors and legacy systems, this pool of AI-adjacent jobs includes positions for systems engineers and software development engineers.
GPT Skill Infrastructure for AI: A US-China Comparison
At present, the United States is better positioned than China to develop the skill infrastructure suitable for AI. First, the United States has more favorable conditions for expanding the number of AI engineers. According to three separate projects that mapped out the global AI talent landscape, many more AI engineers work in the United States than in any other country.94 In 2017, Tencent Research Institute and BOSS Zhipin (a Chinese online job search platform) found that the number of AI “practitioners” (从业者) in the United States far surpassed the corresponding Chinese figure. Figure 7.2 captures this gap across four key AI subdomains: natural language processing (by three times), chips and processors (by fourteen times), machine learning applications (by two times), and computer vision (by three times).95 Overall, the total number of AI practitioners in the United States was two times greater than the corresponding figure for China.96 Furthermore, data from two separate reports by LinkedIn and SCMP Research confirm that the United States leads the world in AI engineers.97
In addition to statistics on the AI workforce, the quantity and quality of AI education is another consideration for which country is better positioned to develop GPT skill infrastructure for AI. Again, the United States leads China by a significant margin in terms of universities with faculty who are proficient in AI. In 2017, the United States was home to nearly half of the world’s 367 universities that provide AI education, operationalized by universities that have at least one faculty member who has published at least one paper in a top AI conference.98 In comparison, China had only 20 universities that met this standard. After replicating this methodology for the years 2020–2021, the US advantage is still pronounced, with 159 universities to China’s 29 universities.99
FIGURE 7.2. A US-China Comparison of AI Practitioners in Key Subdomains. Source: Tencent Research Institute and BOSS Zhipin 2017.
These findings contradict prominent views on the present global distribution of AI engineering talent. In his best-selling book AI Superpowers: China, Silicon Valley, and the New World Order, Kai-Fu Lee argues that the current AI landscape is shifting from an age of discovery, when the country with the highest-quality AI experts wins out, to an age of implementation, when the country with the largest number of sound AI engineers is advantaged.100 In an age of implementation, Lee concludes, “China will soon match or even overtake the United States in developing and deploying artificial intelligence.”101 Pitted against the statistics from the previous passages, Lee’s evidence for China’s lead in AI implementers is meager. His attention is concentrated on anecdotes about the voracious appetite for learning about AI by Chinese entrepreneurs in Beijing.102 While this analysis benefits from Lee’s experience as CEO of Sinovation Ventures, a venture capital fund that invests in many Chinese AI start-ups, it could also be colored by his personal stake in hyping China’s AI capabilities.
Drawing on Lee’s book, Allison and Schmidt also assert that China is cultivating a broader pool of AI talent than the United States today. Specifically, they point out that China graduates three times as many computer science students as the United States on an annual basis.103 Yet the study on which this figure is based finds that computer science graduates in the United States have much higher levels of computer science skills than their Chinese peers. In fact, the average fourth-year computer science undergraduate in the United States scores higher than seniors from the top programs in China.104 Therefore, estimates of China’s pool of AI engineering talent will be misleading if they do not establish some baseline level of education quality. This is another reason to favor the indicators that support an enduring US advantage in AI engineers.105
Second, as previous industrial revolutions have demonstrated, strong linkages between entrepreneurs and scientists that systematize the engineering knowledge related to a GPT are essential to GPT skill infrastructure. In the AI domain, an initial evaluation suggests that this connective tissue is especially strong in the United States. Based on 2015–2019 data, it led the world with the highest number of academic-corporate hybrid AI publications—defined as those coauthored by at least one researcher from both industry and academia—more than doubling the number of such publications from China.106 Xinhua News Agency, China’s most influential media organization, has pinpointed the lack of technical exchanges between academia and industry as one of five main shortcomings in China’s AI talent ecosystem.107
These preliminary indicators align with assessments of the overall state of industry-academia exchanges in China. Barriers to stronger industry-academia linkages include low mobility between institutions, aimless government-sponsored research collaborations, and misguided evaluation incentives for academic researchers.108 One indicator of this shortcoming is the share of R&D outsourced by Chinese firms to domestic research institutes: this figure declined from 2.4 percent in 2010 to 1.9 percent in 2020. Over the same time period, the share of Chinese firms’ R&D expenditures performed by domestic higher education institutions also decreased from 1.2 percent to 0.4 percent.109
Moreover, the US approach to AI standard-setting could prove more optimal for coordinating information flows between labs working on fundamental AI advances and specific application sectors. Market-mediated, decentralized standardization systems are particularly suited for advancing technological domains characterized by significant uncertainty about future trajectories, which clearly applies to AI.110 In such fields, governments confront a “blind giant’s quandary” when attempting to influence technological development through standards-setting.111 The period when government involvement can exert the most influence over the trajectory of an emerging technology coincides with the time when the government possesses the least technical knowledge about the technology. Government intervention therefore could lock in inferior AI standards compared with market-driven standardization efforts.
In that light, China’s state-led approach to technical standards development could hinder the sustainable penetration of AI throughout its economy. For example, the Chinese central government plays a dominant role in China’s AI Industry Alliance, which has pushed to wrest leadership of standards setting in some AI applications away from industry-led standardization efforts.112 Excessive government intervention has been a long-standing weakness of China’s standardization system, producing standards not attuned to market demands and bureaucratic rivalries that undermine the convergence of standards.113 Wang Ping, a leading authority on this topic, has argued that China needs to reform its standardization system to allow private standards development organizations more space to operate, like the Institute of Electrical and Electronics Engineers in the United States and the European Committee for Electrotechnical Standardization.114
In sum, the United States is better positioned than China to not only broaden its pool of AI engineering skills but also benefit from academia-industry linkages in AI engineering. In previous industrial revolutions, these types of institutional adaptations proved crucial to technological leadership. Still, much uncertainty remains in forecasts about GPT skill infrastructure for AI, especially with regard to determining the right measures for the right AI talent. Recent studies of market demand for AI- and ICT-related jobs suggest that employers are softening their demands for a four-year degree in computer science as a requirement for such positions.115 Certificate programs in data science and machine learning that operate under the bachelor-degree level could play an important role in expanding the pool of AI engineering talent.116 Taking into account these caveats, this section’s evaluation of GPT skill infrastructure at the very least calls into question sweeping claims that China is best placed to capitalize on the IR-4.
Reframing National AI Strategies
The preceding conclusions offer a marked contrast with how American and Chinese policymakers devise national AI strategies. Policy proposals for pursuing US leadership in AI consistently call for more AI R&D as the highest priority. For example, the report “Meeting the China Challenge: A New American Strategy for Technology Competition,” published in 2020 by a working group of twenty-eight China specialists and experts, provided sixteen policy recommendations for how the United States should ensure its leadership in AI and three other key technological domains. The very first recommendation was for the United States to significantly expand investment in basic research, raising total R&D funding to at least 3 percent of GDP.117 The Trump administration’s “American AI Initiative,” launched to maintain US leadership in AI “in a time of global power competition,” also listed AI R&D spending as its very first policy recommendation.118
The Chinese government also prioritizes investments in R&D, sometimes at the expense of other routes to productivity growth oriented around technology adoption and education.119 China’s five-year plan (2021–2025) aims to raise basic research spending by over 10 percent in 2021, targeting AI and six other key technological domains.120 China consistently sets and meets ambitious targets for R&D spending, but that same commitment has not extended to education funding. While China’s R&D spending as a percentage of GDP in 2018 was higher than that of Brazil, Malaysia, Mexico, or South Africa (other middle-income countries that industrialized on a similar timeline), China’s public expenditure on education as a percentage of GDP was lower than the figure in those countries.121 According to a group of experts on China’s science and technology policy, one possible explanation for this disparity between attention to R&D versus education is the longer time required for efforts in the latter to yield tangible progress in technological development.122
As both the United States and China transition from initiating a new GPT trajectory to diffusing one across varied application sectors, investing in the broader AI-adjacent skill base will become more crucial than cornering the best and the brightest AI experts. Policies directed at widening the AI talent base, such as enhancing the role of community colleges in developing the AI workforce, deserve more attention.123 Applied technology centers, dedicated field services, and other technology diffusion institutions can incentivize and aid adoption of AI techniques by small and medium-sized enterprises.124 Reorienting engineering education toward maintaining and overseeing AI systems, not solely inventing new ones, also fits this frame.125
A strategy oriented around GPT diffusion does not necessarily exclude support for the exciting research progress in a country’s leading labs and universities. R&D spending undoubtedly will not just help cultivate novel AI breakthroughs but also contribute to widening the GPT skill infrastructure in AI. All too often, however, boosting R&D spending seems to be the boilerplate recommendation for any strategic technology.126 GPTs like AI are not like other technologies, and they demand a different toolkit of strategies.
Alternative Factors
In exploring how the IR-4 could bring about an economic power transition, it is important to compare the implications derived from the GPT diffusion mechanism to those that follow from explanations stressing other factors. Consistent with the previous chapters, I first consider threat-based explanations and the varieties of capitalism (VoC) approach. I then address how US-China competition over emerging technologies could be shaped by differences in regime type, a factor that is particularly salient in this case.
Threat-Based Explanations
One potentially dangerous implication of threat-based explanations is that war, or manufacturing the threat of war, is necessary for economic leadership in the IR-4. Crediting the US military’s key role in spurring advances in GPTs during the twentieth century, Ruttan doubts that the United States could initiate the development of GPTs “in the absence of at least a threat of major war.”127 Extending Ruttan’s line of thinking to the US strategic context in 2014, Linda Weiss expressed concerns that the end of the Cold War, along with the lack of an existential threat, removed the impetus for continued scientific and technological innovation. Specifically, she questioned “why China has not yet metamorphosed into a rival that spurs innovation like the Soviet Union and Japan.”128 Weiss only needed a little more patience. A few years later, the narrative of a US-China “Tech Cold War” gained momentum as both sides of the bilateral relationship trumped up threats to push national scientific and technological priorities.129
GPT diffusion theory strongly refutes the notion that manufacturing external threats is necessary for the United States or China to prevail in the IR-4. An external menace did not drive the rise of the United States in the IR-2. Across all cases, military actors were involved in but not indispensable to spurring the development of GPTs, as many civilian entities also fulfilled the purported role of military investment in providing a large initial demand for incubating GPTs. Furthermore, threat-based interpretations extend only to the moment when one country stimulates the first breakthroughs in a GPT. Even if stoking fears can galvanize support for grand moonshot projects, these do not determine which country is able to benefit most from the widespread adoption of advances in GPTs like AI. That hinges on the more low-key toil of broadening the engineering skill base and advancing interoperability standards in GPTs—not fearmongering.
VoC Explanations
Applying the VoC framework to US-China competition in AI gives more ambiguous results. The VoC approach states that liberal market economies (LME)—prototypically represented by the United States—are more conducive to radical innovation than coordinated market economies (CME).130 It is unclear, however, whether China fits into the VoC framework as a CME or LME. While some label China a CME, others characterize it as an LME.131 This disputed status speaks to the substantial hybridity of China’s economy.132 China has been treated as a “white space on the map” of VoC scholarship, which was originally developed to classify different forms of advanced capitalist economies.133 This makes it difficult to derive strong conclusions from VoC scholarship about China’s ability to adapt to the IR-4’s radical innovations.
The same holds if we focus on the skill formation aspect of the VoC framework. China’s education system emphasizes training for general skills over vocational skills.134 In this respect, it is similar to LMEs like the United States, which means VoC theory provides limited leverage for explaining how the IR-4 could differentially advantage the United States or China. On this topic, GPT diffusion theory points to differences in AI engineering education as more significant than distinctions based on the CME-LME models.
Case-Specific Factors: Regime Type
What is the effect of regime type on technological leadership in the IR-4? The distinction between authoritarian China and the democratic United States takes center stage in arguments about the future of great power rivalry.135 Regime type could also influence the specific aspect of great power competition that GPT diffusion tackles—whether one great power is able to sustain productivity growth at greater rates than its rivals by taking advantage of emerging technologies. Some evidence suggests that, owing to investments in inclusive economic institutions, democracies produce more favorable conditions for growth than autocracies.136 Additionally, empirical work shows that democracies outgrow autocracies in the long run because they are more open to absorbing and diffusing new techniques.137 Other studies find, more specifically, that internet technologies diffuse faster in democracies, possibly because nondemocracies are threatened by the internet’s potential to empower antigovernment movements.138
On the other hand, the impact of democracy on technological progress and economic growth is disputed. Drawing on data from fifty countries over the 1970–2010 period, Taylor finds that regime type does not have a strong relationship with national innovation rates, as measured by patenting rates.139 One review of the econometric evidence on democracy and growth concludes that “the net effect of democracy on growth performance cross-nationally over the last five decades is negative or null.”140 Moreover, China’s rapid economic growth and adoption of internet technologies stands out as an exception to general claims about a democratic advantage when it comes to leveraging new technologies as sources of productivity growth. Contrary to initial expectations, incentives to control online spaces have made some autocratic regimes like China more inclined to spread internet technologies.141 Other scholars point out that the stability of China’s authoritarian regime has encouraged substantial contributions to R&D and technical education, the type of investments in sustained productivity growth typically associated with democracies.142
It is not within the scope of this chapter to settle these debates.143 Still, the juxtaposition of the GPT and LS mechanisms does speak to how regime type could shape US-China competition in the IR-4. Though the conventional wisdom links democracy to freewheeling thought and capacity for innovation, the most important effects of regime type in US-China technological competition during the IR-4, under the GPT diffusion framework, may materialize through changes in GPT skill infrastructure. Democracies tend to be more politically decentralized than autocracies, and decentralized states could be more responsive to the new demands for engineering skills and knowledge in a particular GPT. This accords with evidence that new technologies consistently diffuse more quickly in decentralized states.144
Summary
How far can we take GPT diffusion theory’s implications for the US-China case? I have presented support for the GPT mechanism across a range of historical case studies, each of which covers at least four decades and two countries. At the same time, it is necessary to acknowledge limitations in translating lessons from past industrial revolutions and power transitions to the present.
To begin, it is important to clarify that my findings directly address the mechanism by which technological breakthroughs enable China to surpass the United States in economic productivity.145 The scenario in which China overtakes the United States as the most powerful economy is different from one in which the US-China power gap narrows but does not fully disappear. Scholars have rightly noted that the latter scenario—“posing problems without catching up,” in the words of Thomas Christensen—still significantly bears on issues such as Taiwan’s sovereignty.146 Even so, the possibility of China fully closing the power gap with the United States is especially crucial to study. When rising and established powers are close to parity, according to power transition theory, the risk of hegemonic war is the greatest.147 China’s ability to sustain economic growth also affects its willingness and ability to exert influence in the international arena.
Next, GPT diffusion theory speaks to only one pathway to productivity leadership. The historical case studies have demonstrated that institutional responses to disruptive technological breakthroughs play a key part in economic power transitions. However, China’s prospects for long-term economic growth could also hinge on demographic and geographic drivers.148
A number of factors could affect whether lessons from the past three industrial revolutions extend to the implications of present-day technological advances for a US-China power transition. The most plausible transferability issues can be grouped into those that relate to the nature of great power competition and those that relate to the nature of technological change.
First, as put forward by Stephen Brooks and William Wohlforth, China’s rise in the twenty-first century could face structural barriers that did not exist in previous eras.149 Relying in part on data from 2005–2006, they argue that the present gap between the United States and China in military capabilities, as captured in long-term investments in military R&D, is much larger than past gaps between rising powers and established powers.150 Arguably, the US-China gap in military expenditures has narrowed to the extent that comparisons to historical distributions of military capabilities are more viable. In 2021, China accounted for 14 percent of global military expenditures. This updated figure, albeit still much lower than the US share (38 percent), reflects China’s military modernization efforts over the past two decades during a time of declining US military spending.151 This ratio is more comparable to distributions of military capabilities in the historical periods analyzed in the case studies.152
Another structural barrier China faces is that the growing complexity of developing and deploying advanced military systems now makes it more difficult for rising powers to convert economic capacity into military capacity than it was in the past.153 There are a few reasons why it is still relevant to study China’s ability to convert the technological breakthroughs of the IR-4 into sustained productivity growth. To start, rising states could still benefit from the steady diffusion of some complex military technologies connected to advances in the commercial domain, such as armed uninhabited vehicles.154 In addition, military effectiveness does not solely derive from extremely complex systems like the F-22 stealth fighter. Converting production capacity to military strength could be more relevant for China’s investments in asymmetric capabilities and those suited for specific regional conflicts, such as ground-based air defense systems and the rapid replacement of naval forces.155 Lastly, there remains a strong connection between economic development and countries’ capabilities to “produce, maintain, and coordinate complex military systems.”156
As for the second set of transferability issues, the technological landscape itself is changing. Accelerated globalization of scientific and technological activities may reduce the likelihood of adoption gaps between advanced economies when it comes to emerging technologies.157 Despite these considerations, there are also compelling reasons to think that the nature of technological change in this current period only magnifies the importance of GPT diffusion theory. Cross-country studies indicate that while new technologies are spreading between countries faster than ever, they are spreading to all firms within a country at increasingly slower rates. Networks of multinational firms at the global technology frontiers have reduced cross-national lags in the initial adoption of new technologies, but the cross-national lags in the “intensive adoption” of new technologies, as measured by the time between the technologies’ initial adoption to intensive penetration throughout a country, has only grown.158 These trends give more weight to the GPT mechanism.
Finally, even if the rise and fall of great technologies and powers is fundamentally different in the twenty-first century, previous industrial revolutions still exert substantial influence in the minds of academics and policymakers.159 To justify and sustain their agendas, influential figures in both the United States and China still draw on these historical episodes. At the very least, this chapter submits different lessons to be learned from these guiding precedents.
When some of the leading thinkers of our era declare that the AI revolution will be more significant than the industrial revolution, it is difficult to not get caught up in their excitement. Somehow, every generation winds up believing that their lives coincide with a uniquely important period in history. But our present moment might not be so unprecedented. Unpacking how AI could influence a possible US-China power transition in the twenty-first century requires first learning the lessons of GPT diffusion from past industrial revolutions.
8 Conclusion
STUDIES OF HOW TECHNOLOGY interacts with the international landscape often fixate on the most dramatic aspect of technological change—the eureka moment. Consistent with this frame, the standard explanation for the technological causes of economic power transitions emphasizes a rising power’s ability to dominate profits in leading sectors by generating the first implementation of radical inventions. This book draws attention, in contrast, to the often unassuming process by which an innovation spreads throughout an economy. The rate and scope of diffusion is particularly relevant for GPTs—fundamental advances like electricity or AI that have the potential to drive pervasive transformation across many economic sectors.
Based on the process of GPT diffusion, this book puts forward an alternative theory of how and when significant technological breakthroughs generate differential rates of economic growth among great powers. When evaluating how technological revolutions affect economic power transitions, GPTs stand out as historical engines of growth that can provide major boosts to national productivity. Though each is different, GPTs tend to follow a common pattern: after multiple decades of complementary innovations and institutional adaptations, they gradually diffuse across a broad range of industries. Everything, everywhere, but not all at once.
This impact pathway markedly diverges from existing theories based on leading sectors. Akin to a sprint on a narrow lane, great power competition over leading sectors is framed as a race to dominate initial breakthroughs in the early growth periods of new industries. In contrast, GPT diffusion theory proposes that by more effectively adopting GPTs across many application sectors, some great powers can sustain higher levels of productivity growth than their competitors. Like a marathon on a wide road, great power competition over GPTs is a test of endurance.
Disruptive technological advances can bring about economic power transitions because some countries are more successful at GPT diffusion than others. A nation’s effectiveness at adapting to emerging technologies is determined by the fit between its institutions and the demands of those technologies. Thus, if economic power transitions are driven by the GPT trajectory, as opposed to LS product cycles, the institutional adaptations that matter most are those that facilitate information exchanges between the GPT sector and application sectors, in particular the ability of nations to widen the engineering skill base linked to a new GPT.
Three historical case studies, designed and conducted in a way to assess the explanatory power of the GPT mechanism against the LS mechanism, provide support for GPT diffusion theory. The case studies cover periods characterized by both remarkable technological change—the “three great industrial revolutions” in the eyes of some scholars—and major fluctuations in the global balance of economic power.1 Overall, the case study evidence underscores the significance of GPT diffusion as the key pathway by which the technological changes associated with each industrial revolution translated into differential rates of economic growth among the great powers.
In the case of Britain’s rise to economic preeminence during the First Industrial Revolution, expanded uses of iron in machine-making spurred mechanization, the key GPT trajectory. The gradual progression of mechanization aligned with the period when Britain’s productivity growth outpaced that of France and the Netherlands. Britain’s proficiency in adopting iron machinery across a wide range of economic activities, rather than export advantages from dominating innovation in leading sectors such as cotton textiles, proved more central to its industrial ascent. Though its industrial rivals boasted superior systems of higher technical education for training expert scientists and top-flight engineers, Britain benefited from mechanics’ institutes, educational centers like the Manchester College of Arts and Sciences, and other associations that expanded access to technical literacy and applied mechanics knowledge.
The Second Industrial Revolution case also favors the GPT mechanism’s explanation of why certain great powers better adapt to periods of remarkable technological change. The LS mechanism focuses on Germany’s discoveries in new science-based industries, such as chemicals, as the driving force behind its catching up to Britain before World War I. However, the United States, emerging as the preeminent economic power during this period, was more successful than Germany in exploiting the technological opportunities of the Second Industrial Revolution. Enabled by innovations in machine tools, the extension of interchangeable manufacturing techniques across many American industries functioned as the key GPT trajectory that fueled the rise of the United States. Scientific infrastructure or industrial R&D capabilities, areas in which the United States lagged behind its industrial rivals, cannot account for its advantage in adopting special-purpose machinery across nearly all branches of industry. Rather, the United States gained from institutional adaptations to widen the base of mechanical engineering talent, including through the expansion of technical higher education schools and the professionalization of mechanical engineering.
Evidence from the US-Japan rivalry amid the information revolution exposes more gaps in the LS account. During the late twentieth century Japan captured global market shares in new fast-growing sectors such as consumer electronics and semiconductor components, prompting many to predict that it would overtake the United States as the leading economic power. Yet such an economic power transition, an inevitability based on the expectations of the LS mechanism, never occurred. Instead, the United States sustained higher rates of economic growth than Japan owing, in part, to greater spread of computerization across many economic sectors. Japan’s productivity growth kept up with the US rate in sectors that produced information technology but lagged far behind in sectors that intensively used information technology. Once again, differences in institutional adaptations to widen the GPT skill base turned out to be significant. While Japanese universities were very slow to adapt their training to the demand for more software engineers, US institutions effectively broadened the pool of such skills by cultivating a separate discipline of computer science.
As a supplement to the case studies, I conducted a large-n statistical analysis to test whether countries with superior GPT skill infrastructure preside over higher rates of GPT diffusion. Leveraging time-series cross-sectional data on software engineering education and computerization rates in nineteen countries (the G20 economies) across twenty-five years, the quantitative analysis confirmed this crucial expectation derived from GPT diffusion theory. I found less support for other factors often assumed to have a positive effect on an economy’s broader technological transformation, including institutional factors linked to securing LS product cycles. This empirical test validates a core component of GPT diffusion theory across a sample of the world’s major emerging and developed economies.
Main Contributions
First, at its core, Technology and the Rise of Great Powers introduces and defends GPT diffusion theory as a novel explanation for how and when technological change can lead to a power transition. Historical case studies and statistical analysis substantiate the explanatory power of GPT diffusion theory over the standard explanation of technology-driven power transitions based on leading sectors, which exerts enduring influence in policy and academic circles.2 In doing so, the book answers the call by scholars such as Michael Beckley and Matthew Kroenig for the international relations field to devote more attention to the causes of power transitions, not just their consequences.3
By expounding on the significance of GPT skill infrastructure, the book points toward next steps to better understanding the politics behind some of the most significant technological advances in human history. To investigate why some countries are more successful in cultivating GPT skill infrastructure, promising avenues of research could tap into existing work that concentrates on centralization, inclusiveness of political institutions, government capacity to adopt long time horizons, and industrial organization.4 In these efforts, being careful to differentiate between various pathways by which technological changes make their mark, as this book does with the GPT and LS mechanisms, will be especially important when underlying political factors that satisfy the demands of one type of technological trajectory run counter to the demands of another.
Future work should also probe other institutional factors beyond GPT skill infrastructure that contribute to cross-national differences in GPT adoption. This opens up a universe of institutions that are often ignored in innovation-centered accounts of technological leadership, including gender gaps in engineering education,5 transnational ethnic networks that facilitate technology transfer,6 and “technology diffusion institutions,” such as standard-setting organizations and applied technology centers.7
In positioning the LS mechanism as the main foil to the GPT mechanism, my intention is to use this clash between theories to productively advance our understanding of the rise and fall of great technologies and powers. Contestation should not be misread as disparagement. In one sense, GPT diffusion theory builds on previous scholarship about leading sectors, which first identified the need to flesh out more specific linkages between certain technological advances and more highly aggregated economic changes in the context of power transitions.8 Testing, revising, and improving upon established theories is essential to gradual yet impactful scientific progress—not so unlike the incremental, protracted advance of a GPT.
Second, the book’s central argument also suggests revisions to assessments of power in international politics. Recognizing that scientific and technological capabilities are becoming increasingly central to a nation’s overall power, researchers tend to equate technological leadership with a country’s ability to initiate “key ‘leading sectors’ that are most likely to dominate the world economy into the twenty-first century.”9 For instance, an influential RAND report, “Measuring National Power in the Postindustrial Age,” proposes a template for measuring national power based on a country’s capacity to dominate innovation cycles in “leading sectors.”10 In this effort, the authors draw directly from LS-based scholarship: “The conceptual underpinnings of this template are inspired by the work of Schumpeter, Rostow, Gilpin, Kennedy, and Modelski and Thompson.”11 This study has gained considerable traction in academic and policymaking circles, inspired further workshops on national power, and has been called “the definitive US study on CNP [Comprehensive National Power].”12
Contrary to these approaches, this book submits that evaluations of scientific and technological power should take diffusion seriously. Assessments that solely rely on indicators of innovation capacity in leading sectors will be misleading, especially if a state lags behind in its ability to spread and embed innovations across productive processes. A more balanced judgement of a state’s potential for technological leadership requires looking beyond multinational corporations, innovation clusters like Silicon Valley, and eye-popping R&D numbers to the humble undertaking of diffusion. It shines the spotlight on a different cast of characters: medium-sized firms in small towns, engineers who tweak and implement new methods, and channels that connect the technological frontier with the rest of the economy.
In an article published in the Review of International Political Economy journal, I illustrated the value of this diffusion-oriented approach in gauging China’s scientific and technological capabilities.13 Preoccupied with China’s growing strength in developing new-to-the-world advances, existing scholarship warns that China is poised to overtake the United States in technological leadership. This is mistaken. There is still a large gap between the United States and China when it comes to the countries’ readiness to effectively spread and utilize cutting-edge technologies, as measured by penetration rates of digital technologies such as cloud computing, smart sensors, and key industrial software. When the focus shifts away from impressive and flashy R&D achievements and highly cited publications, China’s “diffusion deficit” comes to light. Indeed, a diffusion-centric assessment indicates that China is much less likely to become a scientific and technological superpower than innovation-centric assessments predict.
Relatedly, the GPT diffusion framework can be fruitfully applied to debates about the effects of emerging technologies on military power. Major theories of military innovation focus on relatively narrow technological developments, such as aircraft carriers, but the most consequential military implications of technological change might come from more fundamental advances like GPTs. In an article that employs evidence from electricity’s impact on military effectiveness to analyze how AI could affect the future of warfare, Allan Dafoe and I challenge studies that predict AI will rapidly spread to militaries around the world and narrow gaps in capabilities.14
Third, as chapter 7 spells out in detail, GPT diffusion theory provides an alternative model for how revolutionary technologies, in particular AI, could affect the US-China power balance. This, in turn, implies different optimal policies for securing technological advantage. Drawing on the LS template, influential thinkers and policymakers in both the United States and China place undue emphasis on three points: the rapid timeframe of economic payoffs from AI and other emerging technologies; where the initial, fundamental innovations in such technologies cluster; and growth driven by a narrow range of economic sectors.
GPT diffusion theory suggests diverging conclusions on all three dimensions. The key technological trajectory is the relative success of the United States and China in adopting AI advances across many industries in a gradual process that will play out over multiple decades. It will be infeasible for one side to cut the other off from foundational innovations in GPTs. The most important institutional factors, therefore, are not R&D infrastructure or training grounds for elite AI scientists but rather those factors that widen the skill base in AI and enmesh AI engineers in cross-cutting networks with entrepreneurs and scientists.15
Yet, the United States is fixated on dominating innovation cycles in leading sectors. When it comes to their grand AI strategy, US policymakers are engrossed in ensuring that leading-edge innovations do not leak to China, whether by restricting the immigration of Chinese graduate students in advanced technical fields or by imposing export controls on high-end chips for training large models like GPT-3 and ChatGPT.16 A strategy informed by GPT diffusion theory would, instead, prioritize improving and sustaining the rate at which AI becomes embedded in a wide range of productive processes. For instance, in their analysis of almost 900,000 associate’s degree programs, Center for Security and Emerging Technology researchers Diana Gehlhaus and Luke Koslosky identified investment in community and technical colleges as a way to unlock “latent potential” in the US AI talent pipeline.17 This recommendation accords with an OECD working paper on the beneficial effects of a wider ICT skills pool on digital adoption rates across twenty-five European countries. The study finds that “the marginal benefit of training for adoption is found to be twice as large for low-skilled than for high-skilled workers, suggesting that measures that encourage the training of low-skilled workers are likely to entail a double dividend for productivity and inclusiveness.”18
At the broadest level, this book demonstrates a method to unpack the causal effects of technological change on international politics. International relations scholars persistently appeal for the discipline to better anticipate the consequences of scientific and technological change, yet these demands remain unmet. By one measure, between 1990 and 2007, only 0.7 percent of the twenty-one thousand articles published in major international relations journals explicitly dealt with the topic of science and technology.19 One bottleneck to researching this topic, which Harold Sprout articulated back in 1963, is that most theories either grossly underestimate the implications of technological advances or assume that technological advance is the “master variable” of international politics.20
This book shows that the middle ground can be a place for fruitful inquiry. Technology does not determine the rise and fall of great powers, but some technological trends, like the diffusion of GPTs, do seem to possess momentum of their own. Social and political factors, as represented by GPT skill infrastructure, shape the pace and direction of these technological trajectories. This approach is particularly useful for understanding the effects of technological change across larger scales of time and space.21
如前所述,对宪法规范力实现的可能性和局限性的分析,仅能源于对法律上的宪法的实效性的深刻把握。这绝非前人未见之论。这一点对于立宪主义国家理论而言是不言而喻的,将宪法从国家现实的整体中分离出来的观念对其而言也是匪夷所思的。这一点在威廉·洪堡(Wilhelm von Humboldt)的政治著作中有着最明确的表述。
康拉德·黑塞(Konrad Hesse, 1919—2005)。刘亚巍、曾韬 译,本文德语版原载K. Hesse, Die Normative Kraft der Verfassung(1959),J. C. B.Mohr(Paul Siebeck),Tübingen,为黑塞1958年在弗莱堡大学法律系的就职演讲稿。
在智利,丢手绢叫“Corre, Corre la Guaraca(快跑快跑小傻瓜)”,玩法跟我们大同小异;“123不许动”在希腊叫“我是一座雕像”,区别在于他们可能带了点cosplay的成分;“石头剪子布”在苏门答腊群岛叫“蚂蚁大象人”。抛羊拐在韩国抛的是石头,在东南亚抛的是小沙包,但游戏规则近乎一致。
事实上,法理学界所用的法学实证主义、实证主义法学、法律实证主义,应是对英文positive law或者legal positivism的翻译有些问题,更准确的含义可能是存在法、实在法、成文法,或法律存在/实在/成文主义。其本来的含义,大概是法律是“制定(laid down and set firmly)”或“存在(exists)”的法。(76)然而,实证主义法学从实证角度理解法,反而在概念上阴差阳错地走向了以实证为核心的实证法学道路。其最大的贡献,是讲明了法律的起源和法律的内容。例如,博登海默在评价奥斯汀的positive law时指出,“奥斯汀希望将普通法排除在成文法之外,因为普通法并不能归结为是君主的命令”。(77)也正因为这样,奥斯汀所提出的法律是由一个君主(或他的代理人)所发出的命令,才招致legal positivism是关于“独裁”的批评。(78)
自苏力提出社科法学后,有学者尝试解读社科法学。例如,王夏昊解读为“社科法学是指以其他社会科学的方法研究法律的学科的总称”。(94)更具体的内容,大概在2014年前后,才由苏力或侯猛单独或一起总结完成。在《法律与社会科学》2014年8月出版的年刊上,徐涤宇在“什么是社科法学”的框架下,提出“首先要讨论的问题是社科法学到底是什么”。侯猛据此正式解读出与王夏昊相似的社科法学的概念,但他的贡献是放在英语世界来理解这个含义。侯猛认为,英文概念“Law and Social Science”或“Social Science of Law”比中文概念“一种跨学科的研究或者说跨学科的知识,即法学和其他学科的知识”更加清晰。侯猛进一步强调社科法学的基调是跨学科,并因为对法学的交叉科学、跨学科法律研究、法律和社会科学、跨法学等总感觉不行,以及鉴于更简洁和上口的表达、更容易和法教义学对话等原因,“直接称为社科法学”。(95)
就在苏力发文的同一期学刊中,侯猛再次提出“社科法学的英文名称是Social Science of Law。中文直译‘法律的社会科学’,只是简称社科法学而已”。在文章里,他虽然澄清社科法学不能被“误认为是法学的分支学科”,(99)但在评论“不再是法社会学”时,事实上又在强调“法社会学转向社科法学”。(100)或许,从某种意义上来讲,社科法学可能正是法社会学的代名词。这样的论断从季卫东将社科法学和法社会学交替使用,也可看出其端倪。(101)6年以后,侯猛发文再次更新了社科法学的定义,将其描述为“法社科研究,全称是法律的社会科学研究(social sciences of law),又简称社科法学,是指运用社会科学的知识和方法来研究法律问题”。(102)与之前社科法学处在云雾里不同,侯猛这时的解释应该才是最清晰和直接的。在文章中,侯猛提出了与苏力早先所强调的社科法学的进路有相似之处,但社科研究与实证研究的关系上却有所不同。一方面,他肯定社科法学和实证研究的通用含义,因为社会科学方法也包括定性和定量。另一方面,他又注意到运用自然科学进行法律实证研究不能与法社科研究等同,同时并非所有法社科研究都可称为实证研究,尤其是冯象和苏力的法律与文学作品不再是实证研究。(103)
2012年,在计量方法方面,屈茂辉更加明确地在定量层面强调实证研究方法。所谓计量法学方法,是“实证研究中通过对研究对象的观察、实验和调查会产生大量数据,必须对这些数据进行统计分析,探寻各个影响变量之间复杂的因果联系”。(112)与此同时,他明确提出这种计量法学方法就是必须使用的定量方法,并阐述了计量法学、计量研究、实证研究对民法学研究的重要性。(113)同年,屈茂辉再次在学科概念下,围绕“大样本”全面阐述计量法学。“计量法学是指通过收集大样本数据,对具有数量变化关系的法律现象进行运用定量研究的交叉学科。它是一门独立的学科,其研究对象是具有数量变化关系的法律现象,研究方法是实证方法和计量方法。”屈茂辉阐明,计量法学的英文渊源是Lee Loevinger于1949年发表的Jurimetrics:the Next Step Forward。(114)在学科价值方面,他认为计量法学是对当前法学研究方法的创新,使中国法学向精细化方向发展,是实现中国法学的国际化途径之一。在实践价值方面,他总结了计量法学的三个贡献:计量法学自身确定的客观标准可作为社会控制和监督的工具,运用系统、实证的观测对其他权力的运用方式来实现;计量法学通过得到控制和监督结果进而反思效用,用定量方法来对政府政策绩效进行评估;计量法学改变了传统方式的法学体系,注重引入统计、计量和社会效果的预测评估方法,更加强调法学的定量、实证和技术性。据此,屈茂辉认为,计量法学是颠覆了传统法学的研究方法。(115)
可见,古代西亚地区的“书同文”是真实的,但这种“书同文”只流于表面,背后的实际情况是:在“书同文”过程中,楔形文字经历了三次脱胎换骨的根本变化,第一次变化发生于公元前2400年前后,从表意文字体系发展出音节文字体系(或音节—表意体系);第二次变化发生于公元前14世纪,在音节文字的基础上,地中海沿岸产生楔形字母,即乌迦里特字母(30个辅音符号);第三次变化发生于公元前6世纪的古波斯帝国,在埃兰音节文字的基础上产生古波斯楔形字母+表意字的混合文字体系,36个字母+5个表意字。第一个在波斯波利斯(Persepolis)完整而准确地临摹古波斯语铭文的尼布尔(Karsten Niebuhr,1733—1815)在完全读不懂铭文的情况下,仅凭直觉判断,认为书写古波斯语的楔形文字是字母(Buchstaben)文字。德国的格罗特芬(G. F. Grotefend,1775─1853)是第一个成功解读古波斯语铭文的人,而他是把这种文字当作字母文字来解读的,因而获得成功,例如,他把书写“大流士”的7个符号解读为d─a─r─h─e─u─sh,显然,在格罗特芬看来,这七个符号就是七个字母,代表语音中的最小单位。从20世纪50年代起,学术著作中的古波斯字母表都成了音节表,a、i、u、ka、ku、ga、gu等等。专门研究古波斯语语法的美国宾大教授肯特(R. G. Kent)认为,每个辅音都自带一个“固有”(inherent)的元音。他一边这样认为,一边又将(仅举一例)“我是大流士”音译为adam:Drayavau?,而不是adama:Drayavau?a,这令人费解。依我浅见,古波斯的这套文字体系属于字母+表意字的混合文字体系,36个字母+5个表意字。在36个字母中,除三个元音(a、i、u)字母外,其余都是辅音字母,不自带“固有”的元音,元音需由阅读者根据语言中的正确形式自行添加。很多(如果不是全部的)文字体系,包括这套古波斯文字体系,都是为某种特定语言发明的,更是为以那种特定语言为母语的人发明的。就古波斯的这套字母而言,只要波斯人掌握了这套辅音字母的发音,就能正确地书写和阅读,也就是说,这套字母文字体系具有与生俱来的助记性质,不完全表达语言。
感知机是一种经典的单层神经网络模型,它通过线性阈值学习过程将多元刺激 x 映射到二分类标签 y 上。为了实现这种映射,感知机通过神经网络权重进行线性变换,并且权重会基于真实标签 t 的反馈进行更新。也就是说,感知机只有在出错时才进行学习。自然的,人们会期望最佳学习与最大错误率相关。然而,因为感知机学习规则实际上是基于梯度下降的,所以前面的分析对这里也适用,即训练的最佳错误率应该是15.87%。
这个结果理论在人工神经和类生物学神经网络具有同样的效果。即“85%规则”既适用于包括多层前馈神经网络、递归神经网络、基于反向传播的各种深度学习算法、玻尔兹曼机、甚至水库计算网络(reservoir computing networks)[21, 22])等广泛的机器学习算法。通过对∂ER/∂β梯度最大化的分析,也证明其适用于类生物神经网络的学习,甚至任何影响神经表征精确度的过程,比如注意、投入或更一般的认知控制[23,24]。例如在后者中,当∂ER/∂β最大化时,参与认知控制的好处会最大化。通过关联预期价值控制理论(Expected Value of Control theory)[23,24,25]的研究,可以知道学习梯度 ∂ER/∂β 由大脑中与控制相关的区域 ( 如前扣带回皮层 ) 来进行监控。
例如,在知觉和审美方面的研究中,俄勒冈大学(University of Oregon)的物理学 Richard Taylor 通过对视觉分形图案的研究发现,如设白纸的维度D为1,一张完全涂黑的纸的维度D为2,即画出来的图形维度在 1~2 之间,那么人类的眼睛更偏好于看维度 D=1.3 的图形[26]。事实上许多大自然物体具有的分形维度就是 1.3,在这个复杂度上人们会感到最舒适。一些著名的艺术家,比如抽象表现主义代表人物 ( Jackson Pollock ),他所画的具有分形的抽象画(下图中间一列,左边是自然图,右边为计算机模拟图)分布在 D=1.1 和 1.9 之间,具有更高分形维度的画面会给人带来更大的压迫感[27]。
心理学家 Rolf Reber 在审美愉悦加工的流畅度理论(Processing fluency theory of aesthetic pleasure)中[28]提出,我们有上述这种偏好是因为大脑可以快速加工这些内容。当我们能迅速加工某些内容的时候,就会获得一个正性反应。例如加工 D = 1.3的分形图案时速度很快,所以就会获得愉悦的情绪反应。此外,在设计和艺术领域心理学家域唐纳德·诺曼(Donald Arthur Norman)和艺术史学家贡布里希(Ernst Gombrich)也分别提出过类似思想。
对比下 D = 1.3 和 15.87% 的出错率,如果进行下统一比例,会发现前者多出原有分形维复杂性和整体的配比,未知:已知(或熟悉:意外,秩序与复杂)约为 0.3/1.3 ≈ 23.07%,这个结果比15.87%要大。这种计算方法最早由数学家 George David Birkhoff 在1928 年于《Aesthetic Measure》一书中提出,他认为若 O 为秩序,C 为复杂度,则一个事物的审美度量 M = O/C。
Celeste Kidd, Steven T Piantadosi, and Richard N Aslin. The goldilocks effect: Human infants allocate attention to visual sequences that are neither too simple nor too complex. PloS one, 7(5):e36399, 2012.
Janet Metcalfe. Metacognitive judgments and control of study. Current Directions in Psychological Science, 18(3):159–163, 2009.
BF Skinner. The behavior of organisms: An experimental analysis. new york: D.appleton-century company, 1938.
Douglas H Lawrence. The transfer of a discrimination along a continuum. Journal of Comparative and Physiological Psychology, 45(6):511, 1952.
J L Elman. Learning and development in neural networks: the importance of starting small. Cognition, 48(1):71–99, Jul 1993.
Kai A Krueger and Peter Dayan. Flexible shaping: How learning in small steps helps.Cognition, 110(3):380–394, 2009.
Yoshua Bengio, Jérˆ ome Louradour, Ronan Collobert, and Jason Weston. Curricu- lum learning. In Proceedings of the 26th annual international conference on machine learning, pages 41–48. ACM, 2009.
M Pawan Kumar, Benjamin Packer, and Daphne Koller. Self-paced learning for latent variable models. In Advances in Neural Information Processing Systems, pages 1189–1197, 2010.
David E Rumelhart, Geoffrey E Hinton, Ronald J Williams, et al. Learning represen- tations by back-propagating errors. Cognitive modeling, 5(3):1, 1988.
Yann LeCun, Yoshua Bengio, and Geoffrey Hinton.Deep learning.Nature, 521(7553):436–444, 2015.
Chi-Tat Law and Joshua I Gold. Reinforcement learning can account for associative and perceptual learning on a visual-decision task. Nat Neurosci, 12(5):655–63, May 2009.
WI Schöllhorn, G Mayer-Kress, KM Newell, and M Michelbrink.Time scales of adaptive behavior and motor learning in the presence of stochastic perturbations.Human movement science, 28(3):319–333, 2009.
Ronald J Williams. Simple statistical gradient-following algorithms for connectionist reinforcement learning. Machine learning, 8(3-4):229–256, 1992.
Frank Rosenblatt. The perceptron: A probabilistic model for information storage and organization in the brain. Psychological review, 65(6):386, 1958.
William T Newsome and Edmond B Pare. A selective impairment of motion perception following lesions of the middle temporal visual area (mt). Journal of Neuroscience, 8(6):2201–2211, 1988.
Kenneth H Britten, Michael N Shadlen, William T Newsome, and J Anthony Movshon.The analysis of visual motion: a comparison of neuronal and psychophysical perfor- mance. Journal of Neuroscience, 12(12):4745–4765, 1992.
Mihaly Csikszentmihalyi. Beyond boredom and anxiety. Jossey-Bass, 2000.
Matti Vuorre and Janet Metcalfe. The relation between the sense of agency and the experience of flow. Consciousness and cognition, 43:133–142, 2016.
Robert Bauer, Meike Fels, Vladislav Royter, Valerio Raco, and Alireza Gharabaghi.Closed-loop adaptation of neurofeedback based on mental effort facilitates reinforce- ment learning of brain self-regulation. Clinical Neurophysiology, 127(9):3156–3164, 2016.
De Houwer J1, Barnes-Holmes D, Moors A..What is learning? On the nature and merits of a functional definition of learning.https://www.ncbi.nlm.nih.gov/pubmed/23359420
Herbert Jaeger. The “echo state” approach to analysing and training recurrent neural networks-with an erratum note. Bonn, Germany: German National Research Center for Information Technology GMD Technical Report, 148(34):13, 2001.
Wolfgang Maass, Thomas Natschläger, and Henry Markram. Real-time computing without stable states: A new framework for neural computation based on perturba- tions. Neural computation, 14(11):2531–2560, 2002.
Amitai Shenhav, Matthew M Botvinick, and Jonathan D Cohen. The expected value of control: an integrative theory of anterior cingulate cortex function. Neuron, 79(2):217–240, 2013.
Amitai Shenhav, Sebastian Musslick, Falk Lieder, Wouter Kool, Thomas L Griffiths, Jonathan D Cohen, and Matthew M Botvinick. Toward a rational and mechanistic account of mental effort. Annual Review of Neuroscience, (0), 2017.
Joshua W Brown and Todd S Braver. Learned predictions of error likelihood in the anterior cingulate cortex. Science, 307(5712):1118–1121, 2005.
Hagerhall, C., Purcell, T., and Taylor, R.P. (2004). Fractal dimension of landscape silhouette as a predictor for landscape preference. Journal of Environmental Psychology 24: 247–55.
A Di Ieva.The Fractal Geometry of the Brain.
Rolf Reber, Norbert Schwarz, Piotr Winkielman.Processing Fluency and Aesthetic Pleasure:Is Beauty in the Perceiver’s Processing Experience.http://dx.doi.org/10.1207/s15327957pspr0804_3
Rigau,Jaume Feixas,Miquel Sbert,Mateu.Conceptualizing Birkhoff’s Aesthetic Measure Using Shannon Entropy and Kolmogorov Complexity. https://doi.org/10.2312/COMPAESTH/COMPAESTH07/105-112
Contents 1 Reading the Animal 2 ‘Paintings’ and ‘Statues’ 3 In the Depths of the Palimpsest 4 Reverse Engineering 5 Common Problem, Common Solution 6 Variations on a Theme 7 In Living Memory 8 The Immortal Gene 9 Out Beyond the Body Wall 10 The Backward Gene’s-Eye View 11 More Glances in the Rear-View Mirror 12 Good Companions, Bad Companions 13 Shared Exit to the Future
1 Reading the Animal
You are a book, an unfinished work of literature, an archive of descriptive history. Your body and your genome can be read as a comprehensive dossier on a succession of colorful worlds long vanished, worlds that surrounded your ancestors long gone: a genetic book of the dead. This truth applies to every animal, plant, fungus, bacterium, and archaean but, in order to avoid tiresome repetition, I shall sometimes treat all living creatures as honorary animals. In the same spirit, I treasure a remark by John Maynard Smith when we were together being shown around the Panama jungle by one of the Smithsonian scientists working there: ‘What a pleasure to listen to a man who really loves his animals.’ The ‘animals’ in question were palm trees.
From the animal’s point of view, the genetic book of the dead can also be seen as a predictor of the future, following the reasonable assumption that the future will not be too different from the past. A third way to say it is that the animal, including its genome, embodies a model of past environments, a model that it uses to, in effect, predict the future and so succeed in the game of Darwinism, which is the game of survival and reproduction, or, more precisely, gene survival. The animal’s genome makes a bet that the future will not be too different from the pasts that its ancestors successfully negotiated.
I said that an animal can be read as a book about past worlds, the worlds of its ancestors. Why didn’t I use the present tense: read the animal as a description of the environment in which it itself lives? It can indeed be read in that way. But (with reservations to be discussed) every aspect of an animal’s survival machinery was bequeathed via its genes by ancestral natural selection. So, when we read the animal, we are actually reading past environments. That is why my title includes ‘the dead’. We are talking about reconstructing ancient worlds in which successive ancestors, now long dead, survived to pass on the genes that shape the way we modern animals are. At present it is a difficult undertaking, but a scientist of the future, presented with a hitherto unknown animal, will be able to read its body, and its genes, as a detailed description of the environments in which its ancestors lived.
I shall have frequent recourse to my imagined Scientist Of the Future, confronted with the body of a hitherto unknown animal and tasked with reading it. For brevity, since I’ll need to mention her often, I shall use her initials, SOF. This distantly resonates with the Greek sophos, meaning ‘wise’ or ‘clever’, as in ‘philosophy’, ‘sophisticated’, etc. In order to avoid ungainly pronoun constructions, and as a courtesy, I arbitrarily assume SOF to be female. If I happened to be a female author, I’d reciprocate.
This genetic book of the dead, this ‘readout’ from the animal and its genes, this richly coded description of ancestral environments, must necessarily be a palimpsest. Ancient documents will be partially over-written by superimposed scripts laid down in later times. A palimpsest is defined by the Oxford English Dictionary as ‘a manuscript in which later writing has been superimposed on earlier (effaced) writing’. A dear colleague, the late Bill Hamilton, had the engaging habit of writing postcards as palimpsests, using different-colored inks to reduce confusion. His sister Dr Mary Bliss kindly lent me this example.
Besides his card being a nicely colorful palimpsest, it is fitting to use it because Professor Hamilton is widely regarded as the most distinguished Darwinian of his generation. Robert Trivers, mourning his death, said, ‘He had the most subtle, multi-layered mind I have ever encountered. What he said often had double and even triple meanings so that, while the rest of us speak and think in single notes, he thought in chords.’ Or should that be palimpsests? Anyway, I like to think he would have enjoyed the idea of evolutionary palimpsests. And, indeed, of the genetic book of the dead itself.
Both Bill’s postcards and my evolution palimpsests depart from the strict dictionary definition: earlier writings are not irretrievably effaced. In the genetic book of the dead, they are partially overwritten, still there to be read, albeit we must peer ‘through a glass darkly’, or through a thicket of later writings. The environments described by the genetic book of the dead run the gamut from ancient Precambrian seas, via all intermediates through the mega-years to very recent. Presumably some kind of weighting balances modern scripts versus ancient ones. I don’t think it follows a simple formula like the Koranic rule for handling internal contradictions – new always trumps old. I’ll return to this in Chapter 3.
If you want to succeed in the world you have to predict, or behave as if predicting, what will happen next. All sensible prediction must be based on the past, and much sensible prediction is statistical rather than absolute. Sometimes the prediction is cognitive – ‘I foresee that if I fall over that cliff (seize that snake by its rattling tail, eat those tempting belladonna berries), it is likely that I will suffer or die in consequence.’ We humans are accustomed to predictions of that cognitive kind, but they are not the predictions I have in mind. I shall be more concerned with unconscious, statistical ‘as-if’ predictions of what might affect an animal’s future chances of surviving and passing on copies of its genes.
This horned lizard of the Mojave, whose skin is tinted and patterned to resemble sand and small stones, embodies a prediction, by its genes, that it would find itself born (well, hatched) into a desert. Equivalently, a zoologist presented with the lizard could read its skin as a vivid description of the sand and stones of the desert environment in which its ancestors lived. And now here’s my central message. Much more than skin deep, the whole body through and through, its very warp and woof, every organ, every cell and biochemical process, every smidgen of any animal, including its genome, can be read as describing ancestral worlds. In the lizard’s case it will no doubt spin the same desert yarn as the skin. ‘Desert’ will be written into every reach of the animal, plus a whole lot more information about its ancestral past, information far exceeding what is available to present-day science.
The lizard burst out of the egg endowed with a genetic prediction that it would find itself in a sun-parched world of sand and pebbles. If it were to violate its genetic prediction, say by straying from the desert onto a golf green, a passing raptor would soon pick it off. Or if the world itself changed, such that its genetic predictions turned out to be wrong, it would also likely be doomed. All useful prediction relies on the future being approximately the same as the past, at least in a statistical sense. A world of continual mad caprice, an environmental bedlam that changed randomly and undependably, would render prediction impossible and put survival in jeopardy. Fortunately, the world is conservative, and genes can safely bet on any given place carrying on pretty much as before. On those occasions when it doesn’t – say after a catastrophic flood or volcanic eruption or, as in the case of the dinosaurs’ tragic end when an asteroid-strike ravaged the world – all predictions are wrong, all bets are off, and whole groups of animals go extinct. More usually, we aren’t dealing with such major catastrophes: not huge swathes of the animal kingdom being wiped out at a stroke, but only those variant individuals whose predictions are slightly wrong, or slightly more wrong than those of competitors within their own species. That is natural selection.
The top scripts of the palimpsest are so recent that they are of a special kind, written during the animal’s own lifetime. The genes’ description of ancestral worlds is overlain by modifications and detailed refinements scripted since the animal was born – modifications written or rewritten by the animal’s learning from experience; or by the remarkable memory of past diseases laid down by the immune system; or by physiological acclimatisation, to altitude, say; or even by simulations in imagination of possible future outcomes. These recent palimpsest scripts are not handed down by the genes (though the equipment needed to write them is), but they still amount to information from the past, called into service to predict the future. It’s just that it’s the very recent past, the past enclosed within the animal’s own lifetime. Chapter 7 is about those parts of the palimpsest that were scribbled in since the animal was born.
There is also an even more recent sense in which an animal’s brain sets up a dynamic model of the immediately fluctuating environment, predicting moment to moment changes in real time. Writing this on the Cornish coast, I take envious pleasure in the gulls as they surf the wind battering the cliffs of the Lizard peninsula. The wings, tail, and even head angle of each bird sensitively adjust themselves to the changing gusts and updraughts. Imagine that SOF, our zoologist of the future, implants radio-linked electrodes in a flying gull’s brain. She could obtain a readout of the gull’s muscle-adjustments, which would translate into a running commentary, in real time, on the whirling eddies of the wind: a predictive model in the brain that sensitively fine-tunes the bird’s flight surfaces so as to carry it into the next split second.
I said that an animal is not only a description of the past, not just a prediction of the future, but also a model. What is a model? A contour map is a model of a country, a model from which you can reconstruct the landscape and navigate its byways. So too is a list of zeros and ones in a computer, being a digitised rendering of the map, perhaps including information tied to it: local population size, crops grown, dominant religions, and so on. As an engineer might understand the word, any two systems are ‘models’ of each other if their behavior shares the same underlying mathematics. You can wire up an electronic model of a pendulum. The periodicity of both pendulum and electronic oscillator are governed by the same equation. It’s just that the symbols in the equation don’t stand for the same things. A mathematician could treat either of them, together with the relevant equation written on paper, as a ‘model’ of any of the others. Weather forecasters construct a dynamic computer model of the world’s weather, continually updated by information from strategically placed thermometers, barometers, anemometers, and nowadays above all, satellites. The model is run on into the future to construct a forecast for any chosen region of the world.
Sense organs do not faithfully project a movie of the outer world into a little cinema in the brain. The brain constructs a virtual reality (VR) model of the real world outside, a model that is continuously updated via the sense organs. Just as weather forecasters run their computer model of the world’s weather into the future, so every animal does the same thing from second to second with its own world model, in order to guide its next action. Each species sets up its own world model, which takes a form useful for the species’ way of life, useful for making vital predictions of how to survive. The model must be very different from species to species. The model in the head of a swallow or a bat must approximate a three-dimensional, aerial world of fast-moving targets. It may not matter that the model is updated by nerve impulses from the eyes in the one case, from the ears in the other. Nerve impulses are nerve impulses are nerve impulses, whatever their origin. A squirrel’s brain must run a VR model similar to that of a squirrel monkey. Both have to navigate a three-dimensional maze of tree trunks and branches. A cow’s model is simpler and closer to two dimensions. A frog doesn’t model a scene as we would understand the word. The frog’s eye largely confines itself to reporting small moving objects to the brain. Such a report typically initiates a stereotyped sequence of events: turning towards the object, hopping to get nearer, and finally shooting the tongue towards the target. The eye’s wiring-up embodies a prediction that, were the frog to shoot out its tongue in the indicated direction, it would be likely to hit food.
My Cornish grandfather was employed by the Marconi company in its pioneering days to teach the principles of radio to young engineers entering the company. Among his teaching aids was a clothesline that he waggled as a model of sound waves – or radio waves, for the same model applied to both, and that’s the point. Any complicated pattern of waves – sound waves, radio waves, or even sea waves at a pinch – can be broken down into component sine waves – ‘Fourier analysis’, named after the French mathematician Joseph Fourier (1768–1830). These in turn can be summed again to reconstitute the original complex wave (Fourier synthesis). To demonstrate this, Grandfather attached his clothesline to rotating wheels. When only one wheel turned, the rope executed serpentine undulations approximating a sine wave. When a coupled wheel rotated at the same time, the rope’s snaking waves became more complex. The sum of the sine waves was an elementary but vivid demonstration of the Fourier principle. Grandfather’s snaking rope was a model of a radio wave travelling from transmitter to receiver. Or of a sound wave entering the ear: a compound wave upon which the brain presumably performs something equivalent to Fourier analysis when it unravels, for example, a pattern even as complex as whispered speech plus intrusive coughing against the background of an orchestral concert. Amazingly, the human ear, well, actually, the human brain, can pick out here an oboe, there a French horn, from the compound waveform of the whole orchestra.
Today’s equivalent of my grandfather would use a computer screen instead of a clothesline, displaying first a simple sine wave, then another sine wave of different frequency, then adding the two together to generate a more complex wiggly line, and so on. The following is a picture of the sound waveform – high-frequency air pressure changes – when I uttered a single English word. If you knew how to analyse it, the numerical data embodied in (a much-expanded image of) the picture would yield a readout of what I said. In fact, it would require a great deal of mathematical wizardry and computer power for you to decipher it. But let the same wiggly line be the groove in which an old-fashioned gramophone needle sits. The resulting waves of changing air pressure would bombard your eardrums and be transduced to pulse patterns in nerve cells connected to your brain. Your brain would then without difficulty, in real time, perform the necessary mathematical wizardry to recognise the spoken word ‘sisters’.
Our sound-processing brain software effortlessly recognises the spoken word, but our sight-processing software has extreme difficulty deciphering it when confronted with a wavy line on paper, on a computer screen, or with the numbers that composed that wavy line. Nevertheless, all the information is contained in the numbers, no matter how they are represented. To decipher it, we’d need to do the mathematics explicitly with the aid of a high-speed computer, and it would be a difficult calculation. Yet our brains find it a doddle if presented with the same data in the form of sound waves. This is a parable to drive home the point – pivotal to my purpose, which is why I said it twice – that some parts of an animal are hugely harder to ‘read’ than others. The patterning on our Mojave lizard’s back was easy: equivalent to hearing ‘sisters’. Obviously, this animal’s ancestors survived in a stony desert. But let us not shrink from the difficult readings – the cellular chemistry of the liver, say. That might be difficult in the same way as seeing the waveform of ‘sisters’ on an oscilloscope screen is difficult. But nothing negates the main point, which is that the information, however hard to decipher, is lurking within. The genetic book of the dead may turn out to be as inscrutable as Linear A or the Indus Valley script. But the information, I believe, is all there.
The pattern to the right is a QR code. It contains a concealed message that your human eye cannot read. But your smartphone can instantly decipher it and reveal a line from my favourite poet. The genetic book of the dead is a palimpsest of messages about ancestral worlds, concealed in an animal’s body and genome. Like QR codes, they mostly cannot be read by the naked eye, but zoologists of the future, armed with advanced computers and other tools of their day, will read them.
To repeat the central point, when we examine an animal there are some cases – the Mojave horned lizard is one – where we can instantly read the embodied description of its ancestral environment, just as our auditory system can instantly decipher the spoken word ‘sisters’. Chapter 2 examines animals who have their ancestral environments almost literally painted on their backs. But mostly we must resort to more indirect and difficult methods in order to extract our readout. Later chapters feel their way towards possible ways of doing this. But in most cases the techniques are not yet properly developed, especially those that involve reading genomes. Part of my purpose is to inspire mathematicians, computer scientists, molecular geneticists, and others better qualified than I am, to develop such methods.
At the outset I need to dispel five possible misunderstandings of the main title, Genetic Book of the Dead. First is the disappointing revelation that I am deferring the task of deciphering much of the book of the dead to the sciences of the future. Nothing much I can do about that. Second, there is little connection, other than a poetic resonance, with the Egyptian Books of the Dead. These were instruction manuals buried with the dead, to help them navigate their way to immortality. An animal’s genome is an instruction manual telling the animal how to navigate through the world, in such a way as to pass the manual (not the body) on into the indefinite future, if not actual immortality.
Third, my title might be misunderstood to be about the fascinating subject of Ancient DNA. The DNA of the long dead – well, not very long, unfortunately – is in some cases available to us, often in disjointed fragments. The Swedish geneticist Svante Pääbo won a Nobel prize for jigsawing the genome of Neanderthal and Denisovan humans, otherwise known only from fossils; in the Denisovan case only three teeth and five bone fragments. Pääbo’s work incidentally shows that Europeans, but not sub-Saharan Africans, are descended from rare cases of interbreeding with Neanderthals. Also, some modern humans, especially Melanesians, can be traced back to interbreeding events with Denisovans. The field of ‘Ancient DNA’ research is now flourishing. The woolly mammoth genome is almost completely known, and there are serious hopes of reviving the species. Other possible ‘resurrections’ might include the dodo, passenger pigeon, great auk, and thylacine (Tasmanian wolf). Unfortunately, sufficient DNA doesn’t last more than a few thousand years at best. In any case, interesting though it is, Ancient DNA is outside the scope of this book.
Fourth, I shall not be dealing with comparisons of DNA sequences in different populations of modern humans and the light that they throw on history, including the waves of human migration that have swept over Earth’s land surface. Tantalisingly, these genetic studies overlap with comparisons between languages. For example, the distribution of both genes and words across the Micronesian islands of the Western Pacific islands shows a mathematically lawful relationship between inter-island distance and word-resemblance. We can picture outrigger canoes scudding across the open Pacific, laden with both genes and words! But that would be a chapter in another book. Might it be called The Selfish Meme?
The present book’s title should not be taken to mean that existing science is ready to translate DNA sequences into descriptions of ancient environments. Nobody can do that, and it’s not clear that SOF will ever do so. This book is about reading the animal itself, its body and behaviour – the ‘phenotype’. It remains true that the descriptive messages from the past are transmitted by DNA. But for the moment we read them indirectly via phenotypes. The easiest, if not the only, way to translate a human genome into a working body is to feed it into a very special interpreting device called a woman.
The Species as Sculpture; the Species as Averaging Computer
Sir D’Arcy Thompson (1860–1948), that immensely learned zoologist, classicist, and mathematician, made a remark that seems trite, even tautological, but it actually provokes thought. ‘Everything is the way it is because it got that way.’ The solar system is the way it is because the laws of physics turned a cloud of gas and dust into a spinning disc, which then condensed to form the sun, plus orbiting bodies rotating in the same plane as each other and in the same direction, marking the plane of the original disc. The moon is the way it is because a titanic bombardment of Earth 4.5 billion years ago hived off into orbit a great quantity of matter, which then was pulled and kneaded by gravity into a sphere. The moon’s initial rotation later slowed, in a phenomenon called ‘tidal locking’, such that we only ever see one face of it. More minor bombardments disfigured the moon’s surface with craters. Earth would be pockmarked in the same way but for erosive and tectonic obliteration. A sculpture is the way it is because a block of Carrara marble received the loving attention of Michelangelo.
Why are our bodies the way they are? Partly, like the moon, we bear the scars of foreign insults – bullet wounds, souvenirs of the duellist’s sabre or the surgeon’s knife, even actual craters from smallpox or chickenpox. But these are superficial details. A body mostly got that way through the processes of embryology and growth. These were, in turn, directed by the DNA in its cells. And how did the DNA get to be the way it is? Here we come to the point. The genome of every individual is a sample of the gene pool of the species. The gene pool got to be the way it is over many generations, partly through random drift, but more pertinently through a process of non-random sculpture. The sculptor is natural selection, carving and whittling the gene pool until it – and the bodies that are its outward and visible manifestation – is the way it is.
Why do I say it’s the species gene pool that is sculpted rather than the individual’s genome? Because, unlike Michelangelo’s marble, the genome of an individual doesn’t change. The individual genome is not the entity that the sculptor carves. Once fertilisation has taken place, the genome remains fixed, from zygote right through embryonic development, to childhood, adulthood, old age. It is the gene pool of the species, not the genome of the individual, that changes under the Darwinian chisel. The change deserves to be called sculpting to the extent that the typical animal form that results is an improvement. Improvement doesn’t have to mean more beautiful like a Rodin or a Praxiteles (though it often is). It means only getting better at surviving and reproducing. Some individuals survive to reproduce. Others die young. Some individuals have lots of mates. Others have none. Some have no children. Others a swarming, healthy brood. Sexual recombination sees to it that the gene pool is stirred and shaken. Mutation sees to it that new genetic variants are fed into the mingling pool. Natural selection and sexual selection see to it that, as generation succeeds generation, the shape of the average genome of the species changes in constructive directions.
Unless we are population geneticists, we don’t see the shifting of the sculpted gene pool directly. Instead, we observe changes in the average bodily form and behaviour of members of the species. Every individual is built by the cooperative enterprise of a sample of genes taken from the current pool. The gene pool of a species is the ever-changing marble upon which the chisels, the fine, sharp, exquisitely delicate, deeply probing chisels of natural selection, go to work.
A geologist looks at a mountain or valley and ‘reads’ it, reconstructs its history from the remote past through to recent times. The natural sculpting of the mountain or valley might begin with a volcano, or tectonic subduction and upthrust. The chisels of wind and rain, rivers and glaciers then take over. When a biologist looks at fossil history, she sees not genes but things that eyes are equipped to see: progressive changes in average phenotype. But the entity being carved by natural selection is the species gene pool.
The existence of sexual reproduction confers on The Species a very special status not shared by other units in the taxonomic hierarchy – genus, family, order, class, etc. Why? Because sexual recombining of genes – shuffling the pack (American deck) – takes place only within the species. That is the very definition of ‘species’. And it leads me to the second metaphor in the title of this section: the species as averaging computer.
The genetic book of the dead is a written description of the world of no particular ancestral individual more than another. It is a description of the environments that sculpted the whole gene pool. Any individual whom we examine today is a sample from the shuffled pack, the shaken and stirred gene pool. And the gene pool in every generation was the result of a statistical process averaged over all those individual successes and failures within the species. The species is an averaging computer. The gene pool is the database upon which it works.
2 ‘Paintings’ and ‘Statues’
When, like that Mojave Desert lizard, an animal has its ancestral home painted on its back, our eyes give us an instant and effortless readout of the worlds of its forebears, and the hazards that they survived. Here’s another highly camouflaged lizard. Can you see it on its background of tree bark? You can, because the photograph was taken in a strong light from close range. You are like a predator who has had the good fortune to stumble upon a victim under ideal seeing conditions. It is such close encounters that exerted the selection pressure to put the finishing touches to the camouflage’s perfection. But how did the evolution of camouflage get its start? Wandering predators, idly scanning out of the corner of their eye, or hunting when the light was poor, supplied the selection pressures that began the process of evolution towards tree bark mimicry, back when the incipient resemblance was only slight. The intermediate stages of camouflage perfection would have relied upon intermediate seeing conditions. There’s a continuous gradient of available conditions, from ‘seen at a distance, in a poor light, out of the corner of the eye, or when not paying attention’ all the way up to ‘close-up, good light, full-frontal’. The lizard of today has a detailed, highly accurate ‘painting’ of tree bark on its back, painted by genes that survived in the gene pool because they produced increasingly accurate pictures.
We have only to glance at this frog to ‘read’ the environment of its ancestors as being rich in grey lichen. Or, in another of Chapter 1’s formulations, the frog’s genes ‘bet’ on lichen. I intend ‘bet’ and ‘read’ in a sense that is close to literal. It requires no sophisticated techniques or apparatus. The zoologist’s eyes are sufficient. And the Darwinian reason for this is that the painting is designed to deceive predatory eyes that work in the same kind of way as the zoologist’s own eyes. Ancestral frogs survived because they successfully deceived predatory eyes similar to the eyes of the zoologist – or of you, vertebrate reader.
In some cases, it is not prey but predators whose outer surface is painted with the colours and patterning of their ancestral world, the better to creep up on prey unseen. A tiger’s genes bet on the tiger being born into a world of light and shade striped by vertical stems. The zoologist examining the body of a snow leopard could bet that its ancestors lived in a mottled world of stones and rocks, perhaps a mountainous region. And its genes place a future bet on the same environment as cover for its offspring.
By the way, the big cat’s mammalian prey might find its camouflage more baffling than we do. We apes and Old World monkeys have trichromatic vision, with three colour-sensitive cell types in our retinas, like modern digital cameras. Most mammals are dichromats: they are what we would call red-green colour-blind. This probably means they’d find a tiger or snow leopard even harder to distinguish from its background than we would. Natural selection has ‘designed’ the stripes of tigers, and the blotches of snow leopards, in such a way as to fool the dichromat eyes of their typical prey. They are pretty good at fooling our trichromat eyes too.
Also in passing, I note how surprising it is that otherwise beautifully camouflaged animals are let down by a dead giveaway – symmetry. The feathers of this owl beautifully imitate tree bark. But the symmetry gives the game away. The camouflage is broken.
I am reduced to suspecting that there must be some deep embryological constraint, making it hard to break away from left-right symmetry. Or does symmetry confer some inscrutable advantage in social encounters? To intimidate rivals, perhaps? Owls can rotate their necks through a far greater angle than we can. Perhaps that mitigates the problem of a symmetrical face. This particular photograph tempts the speculation that natural selection might have favoured the habit of closing one eye because it reduces symmetry. But I suppose that’s too much to hope for.
Subtly different from ‘paintings’ are ‘statues’. Here the animal’s whole body resembles a discrete object that it is not. A tawny frogmouth or a potoo resembling a broken stump of a tree branch, a stick caterpillar sculpted as a twig, a grasshopper resembling a stone or a clod of dry soil, a caterpillar mimicking a bird dropping, are all examples of animal ‘statues’.
The working difference between a ‘painting’ and a ‘statue’ is that a painting, but not a statue, ceases to deceive the moment the animal is removed from its natural background. A ‘painted’ peppered moth removed from the light-coloured bark that it resembles and placed on any other background will instantly be seen and caught by a predator. In this photograph, the background is a soot-blackened tree in an industrial area, which is perfect for the dark, melanic mutant of the same species of moth that you may have noticed less immediately by its side. On the other hand, the masquerading Geometrid stick caterpillar photographed by Anil Kumar Verma in India, if placed on any background, would have a good chance of still being mistaken for a stick and overlooked by a predator. That is the mark of a good animal statue.
Although a statue resembles objects in the natural background, it does not depend for its effectiveness on being seen against that background in the way that a ‘painting’ does. On the contrary, it might be in greater danger. A lone stick insect on a lawn might be overlooked, as a stick that had fallen there. A stick insect surrounded by real sticks might be spotted as the odd one out. When drifting alone, the leafy sea dragon’s resemblance to a wrack might protect it, at least more so than its seahorse cousin whose shape in no way mimics a seaweed. But would this statue be less safe when nestling in a waving bed of real seaweed? It’s a moot question.
Freshwater mussels of the species Lampsilis cardium have larvae that grow by feeding on blood, which they suck from the gills of a fish. The mussel has to find a way to put its larvae into the fish. It does it by means of a ‘statue’, which fools the fish. The mussel has a brood pouch for very young larvae on the edge of its mantle. The brood pouch is an impressive replica of a pair of small fish, complete with false eyes and false, very fish-like, ‘swimming’ movements. Statues don’t move, so the word ‘statue’ is strictly inappropriate, but never mind, you get the point. Larger fish approach and attempt to catch the dummy fish. What they actually catch – and it does them no good – is a squirt of mussel larvae.
This highly camouflaged snake from Iran has a dummy spider at the tip of its tail. It may look only half convincing in a still picture. But the snake moves its tail in such a way that it looks strikingly like a spider scuttling about. Very realistic indeed, especially when the snake itself is concealed in a burrow with only the tail tip visible. Birds swoop down on the spider. And that is the last thing they do. It is worth reflecting on how remarkable it is that such a trick has evolved by natural selection. What might the intermediate stages have looked like? How did the evolutionary sequence get started? I suppose that, before the tip of the tail looked anything like a spider, simply waggling it about was somewhat attractive to birds, who are drawn to any small moving object.
Both ‘paintings’ and ‘statues’ are easy-to-read descriptions of ancestral worlds, the environments in which ancestors survived. The stick caterpillar is a detailed description of ancient twigs. The potoo is a perfect model of long-forgotten stumps. Except that they are not really forgotten. The potoo itself is the memory. Twigs of past ages have carved their own likeness into the masquerading body of that caterpillar. The sands of time have painted their collective self-portrait on the surface of this spider, which you may have trouble spotting.
‘Where are the snows of yesteryear?’ Natural selection has frozen them in the winter plumage of the willow ptarmigan.
The leaf-tailed gecko recalls to our minds, though not his, the dead leaves among which his ancestors lived. He embodies the Darwinian ‘memory’ of generations of leaves that fell long before men arrived in Madagascar to see them, probably long before men existed anywhere.
The green katydid (long-horned grasshopper) has no idea that it embodies a genetic memory of green mosses and fronds over which its ancestors walked. But we can read at a glance that this is so. Same with this adorable little Vietnamese mossy frog.
Statues don’t always copy inanimate objects like sticks or pebbles, dead leaves, or tree branch stubs. Some mimics pretend to be poisonous or distasteful models, and inconspicuous is precisely what they are not. At first glance you might think this was a wasp and hesitate to pick it up. It’s actually a harmless hoverfly. The eyes give it away. Flies have bigger compound eyes than wasps. This feature is probably written in a deep layer of palimpsest that, for some reason, is hard to over-write. The largest anatomical difference between flies and wasps – two wings rather than four (the feature that gives the fly Order its Latin name, Diptera) – is perhaps also difficult to over-write. But maybe, too, that potential clue is hard to notice. What predator is going to take the time to count wings?
Real wasps, the models for the hoverfly mimicry, are not trying to hide. They’re the opposite of camouflaged. Their vividly striped abdomen shouts ‘Beware! Don’t mess with me!’ The hoverfly is shouting the same thing, but it’s a lie. It has no sting and would be good to eat if only the predator dared to attack it. It is a statue, not a painting, because its (fake) warning doesn’t depend on the background. From our point of view in this book, we can read its stripes as telling us that the ecology of its ancestors contained dangerous yellow-and-black stripy things, and predators that feared them. The fly’s stripes are a simulacrum of erstwhile wasp stripes, painted on its abdomen by natural selection. Yellow and black stripes on an insect reliably signify a warning – either true or false – of dire consequences to would-be attackers. The beetle to the right is another, especially vivid example.
If you came face to face with this, peering at you through the undergrowth, would you start back, thinking it was a snake?
It isn’t peering and it isn’t a snake. It’s the chrysalis of a butterfly, Dynastor darius, and chrysalises don’t peer. As a fine pretence of the front end of a snake, it’s well calculated to frighten. Never mind that rational second thoughts could calculate that it’s a bit on the small side to be a dangerous snake. There exists a distance – still close enough to be worrying – at which a snake would look that small. Besides, a panicking bird has no time for second thoughts. One startled squawk and it’s away. Having more time for reflection, the Darwinian student of the genetic book of the dead will read the caterpillar’s ancestral world as inhabited by dangerous snakes. Some caterpillars, whose rear ends pull the same snake trick, even move muscles in such a way that the fake eyes seem to close and open. Would-be predators can’t be expected to know that snakes don’t do that.
Eyes are scary in themselves. That’s why some moths have eyespots on their wings, which they suddenly expose when surprised by a predator. If you had good reason to fear tigers or other members of the cat family, might you not start back in alarm if suddenly confronted with this, the so-called owl moth of South East Asia?
There exists a distance – a dangerous distance – at which a tiger or a leopard would present a retinal image the same size as a close-up moth. OK, it doesn’t look very like any particular member of the cat family to our eyes. But there’s plenty of evidence that animals of various species respond to dummies that bear only a crude resemblance to the real thing – scarecrows are a familiar example, and there’s lots of experimental evidence as well. Black-headed gulls respond to a model gull head on the end of a stick, as though it were a whole real gull. A shocked withdrawal might be all it takes to save this moth.
I am amused to learn that eyes painted on the rumps of cattle are effective in deterring predation by lions.
We could call it the Babar effect, after Jean de Brunhoff’s lovable and wise King of the Elephants, who won the war against the rhinoceroses by painting scary eyes on elephant rumps.
What on Earth is this? A dragon? A nightmare devil horse? It is in fact the caterpillar of an Australian moth, the pink underwing. The spectacular eye and teeth pattern is not visible when the caterpillar is at rest. It is screened by folds of skin. When threatened, the animal pulls back the skin screen to unveil the display, and, well, all I can say is that if I were a would-be predator, I wouldn’t hang about.
PHOTO: HUSEIN LATIF
The scariest false face I know? It’s a toss-up between the octopus on the left and the vulture on the right. The real eyes of the octopus can just be seen above the inner ends of the ‘eyebrows’ of the large, prominent false eyes. You can find the real eyes of the Himalayan griffon vulture if you first locate the beak and hence the real head. The false eyes of the octopus presumably deter predators. The vulture seems to use its false face to intimidate other vultures, thereby clearing a path through a crowd around a carcase.
Some butterflies have a false head at the back of the wings. How might this benefit the insect? Five hypotheses have been proposed, of which the consensus favourite is the deflection hypothesis: birds are thought to peck at the less vulnerable false head, sparing the real one. I slightly prefer a sixth idea, that the predator expects the butterfly to take off in the wrong direction. Why do I prefer it? Perhaps because I am committed to the idea that animals survive by predicting the future.
Paintings and statues aimed at fooling predators constitute the nearest approach achieved by any book of the dead to a literal readout, a literal description of ancestral worlds. And the aspect of this that I want to stress is its astounding accuracy and attention to detail. This leaf insect even has fake blemishes. The stick caterpillar (here) has fake buds.
I see no reason why the same scrupulous attention to detail should not pervade less literal, less obvious parts of the readout. I believe the same detailed perfection is lurking, waiting to be discovered, in internal organs, in brain-wiring of behaviour, in cellular biochemistry, and other more indirect or deeply buried readings that can be dug out if only we could develop the tools to do so. Why should natural selection escalate its vigilance specifically for the external appearance of animals? Internal details, all details, are no less vital to survival. They are equally subject to becoming written descriptions of past worlds, albeit written in a less transparent script, harder to decipher than this chapter’s superficial paintings and statues. The reason paintings and statues are easier for us to read than internal pages of the genetic book of the dead is not far to seek. They are aimed at eyes, especially predatory eyes. And, as already pointed out, predatory eyes, vertebrate ones at least, work in the same way as our eyes. No wonder it is camouflage and other versions of painting and sculpture that most impress us among all the pages of the book of the dead.
I believe the internally buried descriptions of ancestral worlds will turn out to have the same detailed perfection as the externally seen paintings and statues. Why should they not? The descriptions will just be written less literally, more cryptically, and will require more sophisticated decoding. As with the ear’s decoding of Chapter 1’s spoken word ‘sisters’, the paintings and statues of this chapter are effortlessly read pages from books of the dead. But just as the ‘sisters’ waveform, when presented in the recalcitrant form of binary digits, will eventually yield to analysis, so too will the non-obvious, non-skin-deep details of animals and their genes. The book of the dead will be read, even down to minute details buried deep inside every cell.
This is my central message, and it will bear repeating here. The fine-fingered sculpting of natural selection works not just on the external appearance of an animal such as a stick caterpillar, a tree-climbing lizard, a leaf insect or a tawny frogmouth, where we can appreciate it with the naked eye. The Darwinian sculptor’s sharp chisels penetrate every internal cranny and nook of an animal, right down to the sub-microscopic interior of cells and the high-speed chemical wheels that turn therein. Do not be deceived by the extra difficulty of discerning details more deeply buried. There is every reason to suppose that painted lizards or moths, and moulded potoos or caterpillars, are the outward and visible tips of huge, concealed icebergs. Darwin was at his most eloquent in expressing the point.
It may be said that natural selection is daily and hourly scrutinising, throughout the world, every variation, even the slightest; rejecting that which is bad, preserving and adding up all that is good; silently and insensibly working, whenever and wherever opportunity offers, at the improvement of each organic being in relation to its organic and inorganic conditions of life. We see nothing of these slow changes in progress, until the hand of time has marked the long lapse of ages, and then so imperfect is our view into long past geological ages, that we only see that the forms of life are now different from what they formerly were.
3 In the Depths of the Palimpsest
It’s all very well for me to say an animal is a readout of environments from the past, but how far into the past do we go? Every twinge of lower-back pain reminds us that our ancestors only 6 million years ago walked on all fours. Our mammalian spine was built over hundreds of millions of years of horizontal existence when the working body depended on it – depended in the literal sense of hanging from it. The human spine was not ‘meant’ to stand vertically, and it understandably protests. Our human palimpsest has ‘quadruped’ boldly written in a firm hand, then over-written all too superficially – and sometimes painfully – with the tracery of a new description – biped. Parvenu, Johnny-come-lately biped.
The skin of Chapter 1’s Mojave horned lizard proclaimed to us an ancestral world of sandy, stony desert, but that world was presumably recent. What can we read from the palimpsest about earlier environments? Let’s begin by going back a very long way. As with all vertebrates, lizard embryos have gill arches that speak to us of ancestral life in water. As it happens, we have fossils to tell us that the watery scripts of all terrestrial vertebrates, including lizards, date back to Devonian times and then back to life’s marine beginning. The poetic point has often been made – I associate it with that salty, larger-than-life intellectual warrior JBS Haldane – that our saline blood plasma is a relic of Palaeozoic seas. In a 1940 essay called ‘Man as a Sea Beast’, Haldane notes that our plasma is similar in chemical composition to the sea but diluted. He takes this as an indication, not a very strong one in my reluctant opinion (‘reluctant’ because I like the idea), that Palaeozoic seas were less salty than today’s:
As the sea is always receiving salt from the rivers, and only occasionally depositing it in drying lagoons, it becomes saltier from age to age, and our plasma tells us of a time when it possessed less than half its present salt content.
The phrase ‘tells us of a time’ resonates congenially with the title of this book. Haldane goes on:
we pass our first nine months as aquatic animals, suspended in and protected by a salty fluid medium. We begin life as salt-water animals.
Whatever the plausibility of Haldane’s inference about changing salinity, what is undeniable is this. All life began in the sea. The lowest level of palimpsest tells a story of water. After some hundreds of millions of years, plants and then a variety of animals took the enterprising step out onto the land. Following Haldane’s fancy, we could say they eased the journey by taking their private sea water with them in their blood. Animal groups that independently took this step include scorpions, snails, centipedes and millipedes, spiders, crustaceans such as woodlice and land crabs, insects (who later took a further giant leap into the air) and a range of worms who, however, never stray far from moisture to this day. All these animals have ‘dry land’ inscribed on top of the deeper marine layers of palimpsest. Of special interest to us as vertebrates, the lobefins, a group of fish represented today by lungfish and coelacanths, crawled out of the sea, perhaps initially only in search of water elsewhere but eventually to take up permanent residence on dry land, in some cases very dry indeed. Intermediate palimpsest scripts tell of juvenile life in water (think tadpole) accompanying adult emergence on land.
That all makes sense. There was a living to be made on land. The sun showers the land with photons, no less than the surface of the sea. Energy was there for the taking. Why wouldn’t plants take advantage of it via green solar panels, and then animals take advantage of it via plants? Do not suppose that a mutant individual suddenly found itself fully equipped genetically for life on land. More probably, individuals of an enterprising disposition made the first uncomfortable moves. This was perhaps rewarded by a new source of food. We can imagine them learning to make brief, snatch-and-grab forays out of water. Genetic natural selection would have favoured individuals who were especially good at learning the new ploy. Successive generations would have become better and better at learning it, spending less and less time in the sea.
The general name for learned behaviour becoming genetically incorporated is the Baldwin Effect. Though I won’t discuss it further here, I suspect that it’s important in the evolution of major innovations generally, perhaps including the first moves towards defying gravity in flight. In the case of the lobe-finned fishes who left the water in the Devonian era around 400 million years ago, there are various theories for how it happened. One that I like was proposed by the American palaeontologist AS Romer. Recurrent drought would have stranded fishes in shrinking pools. Natural selection favoured individuals able to leave a doomed pool and crawl overland to find another one. A point in strong favour of the theory is that there would have been a continuous range of distances separating the pools. At the beginning of the evolutionary progression, a fish could save its life by crawling to a neighbouring pool only a short distance away. Later in evolution, more distant pools could be reached. All evolutionary advances must be gradual. A suffocating fish’s ability to exploit air requires physiological modification. Major modification cannot happen in one fell swoop. That would be too improbable. There has to be a gradient of step-by-step small improvement. And a gradient of distances between pools, some near, some a bit further, some far, is exactly what is needed. We shall meet the point again in Chapter 6 and the astonishingly rapid evolution of Cichlid fishes in Lake Victoria. Unfortunately, Romer prefaced his theory by quoting evidence that the Devonian was especially prone to drought. When this evidence was called into question, Romer’s whole theory suffered in appreciation. Unnecessarily so.
In whatever way the move to the land happened, profound redesign became necessary. Water really is a very different environment from airy land. For animals, the move out of water was accompanied by radical changes in anatomy and physiology. Watery scripts at the base of the palimpsest had to be comprehensively over-written. It is the more surprising that a large number of animal groups later went into reverse, throwing their hard-won retooling to the winds as they trooped back into the water. Among invertebrates, the list includes pond snails, diving bell spiders, and water beetles. The water that they re-invaded is fresh water, not sea. But some vertebrate returnees, notably whales (including dolphins), sea cows, sea snakes, and turtles, went right back into the salted marine world that their ancestors had taken such trouble to leave.
Seals, sea lions, walruses, and their kin, also Galapagos marine iguanas, only partially returned to the sea, to feed. They still spend much time on land, and breed on land. So do penguins, whose streamlined athleticism in the sea is bought at the cost of risible maladroitness on land. You cannot be a master of all trades. Sea turtles laboriously haul themselves out on land to lay eggs. Otherwise, they totally recommitted to the sea. As soon as baby turtles hatch in the sand, they lose no time in racing down the beach to the sea. Lots of other land vertebrates moved part-time into fresh water, including snakes, crocodiles, hippos, otters, shrews, tenrecs, rodents such as water voles and beavers, desmans (a kind of mole), yapoks (water opossums), and platypuses. These still spend a good deal of time on land, taking to the water mainly to feed.
Sea turtle
You might think that returnees to water would unmask the lower layers of palimpsest and rediscover the designs that served their ancestors so well. Why don’t whales, why don’t dugongs, have gills? Their embryos, like the embryos of all mammals, even have the makings of gills. It would seem the most natural thing in the world to dust off the old script and press it into service again. That doesn’t happen. It’s almost as though, having gone to such trouble to evolve lungs, they were reluctant to abandon them, even if, as you might think, gills would serve them better. Given gills, they wouldn’t have to keep coming to the surface to breathe. But rather than revive the gill, what they did was stick loyally to the lung, even at the cost of profound modifications to the whole air-breathing system, to accommodate the return to water.
They changed their physiology in extreme ways such that they can stay under water for over an hour in some cases. When whales do come to the surface, they can exchange a huge volume of air very quickly in one roaring gulp before submerging again. It’s tempting to toy with the idea of a general rule stating that old scripts from lower down the palimpsest cannot be revived. But I can’t see why this should in general be true. There has to be a more telling reason. I suspect that, having committed their embryological mechanics to air-breathing lungs, the repurposing of gills would be a more radical embryological upheaval, more difficult to achieve than rewriting superficial scripts to modify the air-breathing equipment.
Sea snakes don’t have gills, but they obtain oxygen from water through an exceptionally rich blood supply in the head. Again, they went for a new solution to the problem, rather than revive the old one. Some turtles obtain a certain amount of oxygen from water via the cloaca (waste disposal plus genital opening), but they still have to come to the surface to breathe air into their lungs.
Steller’s sea cow
Never parted from the buoyant support of water, whales are freed to evolve in massively (indeed so) different directions from their terrestrial ancestors. The blue whale is probably the largest animal that ever lived. Steller’s sea cows (see previous page), extinct relatives of dugongs and manatees, reached lengths of 11 metres and masses of 10 tonnes, larger than minke whales. They were hunted to extinction in the eighteenth century, soon after Steller first saw them. Like whales, sea cows breathe air, having failed to rediscover anything equivalent to the gills of their earlier ancestors. For reasons just discussed, that word ‘failed’ may be ill-advised.
Ichthyosaurs were reptilian contemporaries of the dinosaurs, with fins and streamlined bodies, and with powerful tails, which were their main engines of propulsion: like dolphins, except that ichthyosaur tails would have moved from side to side rather than up and down. The ancestors of whales and dolphins had already perfected the mammalian galloping gait on land, and the up-and-down motion of dolphin flukes was naturally derived from it. Dolphins ‘gallop’ through the water, unlike ichthyosaurs, who would have swum more like fish. Otherwise, ichthyosaurs looked like dolphins and they probably lived pretty much like dolphins. Did they leap exuberantly into the air – wonderful thought – wagging their tails like dolphins (but from side to side)? They had big eyes, from which we might guess that they probably didn’t rely on sonar as the small-eyed dolphins do. Ichthyosaurs gave birth to live babies in the sea, as we know from a fossil ichthyosaur who unfortunately died during the act of giving birth (see above). Unlike turtles, but like dolphins and sea cows, ichthyosaurs were fully emancipated from their terrestrial heritage. So were plesiosaurs, for there’s evidence that they were livebearers too. Given that viviparity has evolved, according to one authoritative estimate, at least 100 times independently in land reptiles, it seems surprising that sea turtles, buoyant in water but painfully heavy on land, still labour up the sands to lay eggs. And that their babies, when they hatch, are obliged to flap their perilous way down to the sea, running a gauntlet of gulls, frigate birds, foxes, and even marauding crabs.
Ichthyosaur died while giving birth
Sea turtles revert to land to lay their eggs, in holes that they dig in a sandy beach. And an arduous exertion it is, for they are woefully ill-equipped to move out of water. Seals, sea lions, otters, and many other mammals whom we’ll discuss in a moment, spend part of their time in water and are adapted to swimming rather than walking, which makes them clumsy on land, though less so than sea turtles. As already remarked, the same is true of penguins, who are champions in water but comically awkward on land. Galapagos marine iguanas are proficient swimmers, but they can manage a surprising turn of speed on land too, when fleeing snakes. All these animals show us what the intermediates might have been like, on the way to becoming dedicated mariners like whales, dugongs, plesiosaurs, and ichthyosaurs.
Tortles and turtoises – a tortuous trajectory
Turtles and tortoises are of special interest from the palimpsest point of view, and they deserve special treatment. But first I have to dispel a confusing quirk of the English language. In British common usage, turtles are purely aquatic, tortoises totally terrestrial. Americans call them all turtles, tortoises being those turtles that live on land. In what follows, I’ll try to use unambiguous language that won’t confuse readers from either of the two nations ‘separated by a common language’. I’ll sometimes resort to ‘chelonians’ to refer to the entire group.
Land tortoises, as we shall see, are almost unique in that their palimpsest chronicles a double doubling-back during the long course of their evolution. Their fish ancestors, along with the ancestors of all land vertebrates including us, left the sea in Devonian times, around 400 million years ago. After a period on land they then, like whales and dugongs, like ichthyosaurs and plesiosaurs, returned to the water. They became sea turtles. Finally, uniquely, some aquatic turtles came back to the land and became our modern dry-land (in some cases very dry indeed) tortoises. This is the ‘double doubling-back’ that I mentioned. But how do we know? How has the uniquely complicated palimpsest of land tortoises been deciphered?
We can draw a family tree of extant chelonians, using all available evidence including molecular genetics. The diagram below is adapted from a paper by Walter Joyce and Jacques Gauthier. Aquatic groups are shown in blue, terrestrial in orange. I’ve taken the liberty of colouring the ‘ancestral’ blobs blue when the majority of their descendant groups are blue. Today’s land tortoises constitute a single branch, nested among branches consisting of aquatic turtles.
This suggests that modern land tortoises, unlike most land reptiles and mammals, have not stayed on land continuously since their fish ancestors (who were also ours) emerged from the sea. Land tortoises’ ancestors were among those who, like whales and dugongs, went back to the water. But, unlike whales and dugongs, they then re-emerged back onto the land. I suppose this means I should reluctantly admit that American terminology has something going for it. As it turns out, what we British call tortoises are just sea turtles who turned turtle and returned to the land. They’re terrestrial turtles. No, I can’t do it. My upbringing leads me to go on calling them tortoises, but I’ll curb my tendency to wince at a phrase like ‘desert turtles’. In any case, what is interesting from the point of view of the genetic book of the dead is this: where reversals are concerned, land tortoises appear to have the most complicated palimpsests of all, with the largest number of almost perverse-seeming reversals.
Modern land tortoise
Moreover, it appears that our modern land tortoises may not be the first of their kind to achieve this remarkable double doubling-back. What looks like an earlier case occurred in the Triassic era. Two genera, Proganochelys and Palaeochersis, date way back to the first great age of dinosaurs, indeed long before the more spectacular and famous giant dinosaurs of the Jurassic and Cretaceous. It appears that they lived on land. How can we know? This is a good opportunity to return to our ‘future scientist’ SOF, faced with an unknown animal, and invite her to ‘read’ its environment from its skeleton. Fossils present the challenge in earnest because we can’t watch them living – whether swimming or walking – in their environment.
Proganochelys
So, what might SOF say of those enigmatic fossils, Proganochelys and Palaeochersis? Their feet don’t look like swimming flippers. But can we be more scientific about this? Joyce and Gauthier, whom we’ve already met, used a method that can point the way for anyone who wants to quantitatively decipher the genetic book of the long dead. They took seventy-one living species of chelonians whose habitat is known, and made three key measurements of their arm bones, the humerus (upper arm), the ulna (one of the two forearm bones), and the hand, as a percentage of total arm length. They plotted them on triangular graph paper. Triangular plotting makes convenient use of a proof in Euclidean geometry. From any point inside an equilateral triangle, the lengths of perpendiculars dropped to the three sides add up to the same value. This provides a useful technique for displaying three variables when the three are proportions that add up to a fixed number such as one, or percentages that add up to 100. Each coloured point represents one of the seventy-one species. The perpendicular distances of a point from each of the three lines of the big triangle represent the lengths of their three skeletal measurements. And when you colour-code the species according to whether they live in water or on land, something significant leaps off the page. The coloured points elegantly separate out. Blue points represent species living in water, yellow points species living on land. Green points represent genera that spend time in both environments and they, satisfyingly, occupy the region between the blues and yellows.
So now, the interesting question is, where do the two ancient fossil species, Palaeochersis and Proganochelys, fall? They are represented by the two red stars. And there’s little doubt about it. The red stars fall among the yellow points, the dry-land species of modern tortoises. They were terrestrial tortoises. The two stars fall fairly close to the green points, so maybe they didn’t stray far from water. This kind of method shows one way in which our hypothetical SOF might ‘read’ the environment of any hitherto unknown animal – and hence read the environment in which its ancestors were naturally selected. No doubt SOF will have more advanced methods at her disposal, but studies such as this one might point the way.
Palaeochersis and Proganochelys, then, were landlubbers. But had they stayed on land ever since their (and our) fishy ancestors crawled out of the sea? Or did they, like modern land tortoises, number sea turtles among their forebears? To help decide this, let’s look at another fossil. Odontochelys semitestacea lived in the Triassic, like Palaeochersis and Proganochelys but earlier. It was about half a metre long, including a long tail, which modern chelonians lack. The ‘Odonto’ in the generic name records the fact that it had teeth, unlike all modern chelonians, who have something more like a bird’s beak. And the specific name semitestacea testifies to its having only half a shell. It had a ‘plastron’, the hard shell that protects the belly of all chelonians, but it lacked the domed upper shell. The ribs, however, were flattened like those that support the shell in a normal chelonian.
The fossil was discovered in China and described by a group of scientists led by Li Chun. They believe Odontochelys, or something like it, is ancestral to all chelonians and that the turtle shell evolved ‘from the bottom up’. They referred to the Joyce and Gauthier paper on forelimb proportions and concluded that Odontochelys was aquatic. In case you’re wondering what was the use of half a shell, sharks (who have been around since long before any of this story) often attack from below, so the armoured belly might have been anti-shark. If we accept this interpretation, it again suggests that the chelonian shell evolved in water. Against land predators we would not expect that the breastplate should be the first piece of armour to evolve. Quite the reverse. Odontochelys was probably something like a swimming lizard, a sort of Galapagos marine iguana but armoured with a large ventral breastplate.
Although it’s controversial, the Chinese scientists favour the view that an aquatic turtle like Odontochelys, with its half shell, was ancestral to chelonians. Like all reptiles, it would have been descended from terrestrial, lizard-like ancestors, perhaps something like Pappochelys. If they are right that the chelonian shell evolved, Odontochelys-style, from the bottom up in shark-infested waters, what can we say about Palaeochersis and Proganochelys out on the land?
Odontochelys
It would seem that these represent an earlier emergence from water, an earlier incarnation of doubling-back terrestrial tortoises, to parallel today’s behemoths of Galapagos and Aldabra, who evolved from a later generation of aquatic turtles. In any case, the group we know as land tortoises stand as poster child for the very idea of an elaborate palimpsest. Not only did they leave the water for the land, return to water, and then double back onto the land again. They may even have done it twice! The doubling-back was achieved first by the likes of Proganochelys, and then again, independently, by our modern land tortoises. Maybe some went back to water yet again. It wouldn’t surprise me if some freshwater terrapins represent such a triple reversal, but I know of no evidence. Even one doubling-back is remarkable enough.
Pappochelys
If this giant Galapagos tortoise could sing a Homeric epic of its ancestors, its DNA-scored Odyssey would range from ancient legends of Devonian fishes, through lizard-like creatures roaming Permian lands, back to the sea with Mesozoic turtles, and finally returning to the land a second time. Now that’s what I call a palimpsest!
Giant Galapagos tortoise
Who Sings Loudest
I said in Chapter 1 that the palimpsest chapter would return to the question of the relative balance between recent scripts and ancient ones. It is time to do so. You might conjecture something like the scriptural rule for internal Koranic contradictions: later verses supersede earlier ones. But it’s not as simple as that. In the genetic book of the dead, older scripts of the palimpsest can amount to ‘constraints on perfection’.
Famous cases of evolutionary bad design, such as the vertebrate retina being installed back to front, or the wasteful detour of the laryngeal nerve (see below), can be blamed on historical constraints of this kind.
‘Can you tell me the way to Dublin?’
‘Well, I wouldn’t start from here.’
The joke is familiar to the point of cliché, but it strikes to the heart of our palimpsest priority question. Unlike an engineer who can go back to the drawing board, evolution always has to ‘start from here’, however unfavourable a starting point ‘here’ may be. Imagine what the jet engine would look like if the designer had had to start with a propellor engine on his drawing board, which he then had to modify, step by tinkering step, until it became a jet engine. An engineer starting with the luxury of a clean drawing board would never have designed an eye with the ‘photocells’ facing backwards, and their output ‘wires’ being obliged to travel over the surface of the retina and eventually dive through it in a blind spot on their way to the brain. The blind spot is worryingly large, although we don’t notice it because the brain, in building its constrained virtual reality model of the world, cunningly fills in a plausible replacement for the missing patch on the visual field. I suppose such guesswork could be dangerous if a hazard happened to fall on the blind spot at a crucial moment. But this piece of bad design is buried deep in embryology. To change it in order to make the end product more sensible would require a major upheaval early in the embryonic development of the nervous system. And the earlier in embryology it is, the more radical and difficult to achieve. Even if such an upheaval could at length be achieved, the intermediate evolutionary stages on the way to the ultimate improvement would probably be fatally inferior to the existing arrangement, which works, after all, pretty well. Mutant individuals who began the long trek to ultimate improvement would be out-competed by rivals who coped adequately with the status quo. Indeed, in the hypothetical case of reforming the retina, they would probably be totally blind.
You can call the backwards retina ‘bad design’ if you wish. It’s a legacy of history, a relic, an older palimpsest script partially over-written. Another example is the tail of humans and other apes, prominent in the embryo, shrunk to the coccyx in the adult. Also faintly traced in the palimpsest is our sparse covering of hair. Once useful for heat insulation, it is now reduced to a relic, still retaining its now almost pointless erectile properties in response to cold or emotion.
The recurrent laryngeal nerve in a mammal or a reptile serves the larynx. But instead of going directly to its destination, it shoots straight past the larynx, on its way down the neck into the chest, where it loops around a major artery and then rushes all the way back up the neck to the larynx. If you think of it as design, this is obviously rotten design. The length of the detour in the giant dinosaur Brachiosaurus would have been about 20 metres. In a giraffe it is still impressive, as I witnessed at first hand when, for a Channel Four documentary called Inside Nature’s Giants, I assisted in the dissection of a giraffe, who had unfortunately died in a zoo. Who knows what inefficiencies or outright errors might have resulted from the transmission delay that such a detour must have imposed. But natural selection is not wantonly silly. It wasn’t originally bad design in our fishy ancestors when the nerve in question went straight to its end organ – not larynx, for fish don’t have a larynx. Fish don’t have a neck either. When the neck started to lengthen in their land-dwelling descendants, the marginal cost of each small lengthening of the detour was small compared to what would have been the major cost of radically reforming embryology to re-route the nerve along a ‘sensible’ path, the other side of the artery. Mutant individuals who began the embryologically radical evolutionary journey towards re-routing the laryngeal nerve would have been out-competed by rival individuals who made do with the working status quo. There’s a very similar example in the routing of the tube connecting testis to penis. Instead of taking the most direct route, it loops over the tube connecting kidney to bladder: an apparently pointless detour. Once again, the bad design is a constraint buried deep in embryology and deep in history.
Recurrent laryngeal nerve
‘Buried deep in embryology and deep in history’ is another way of saying ‘buried deep under layers of younger scripts in the palimpsest’. Far from a ‘Koranic’ type of rule in which ‘Later trumps Earlier’, we might be tempted to toy with the reverse, ‘Earlier trumps Later’. But that won’t do either. The selection pressures that winnowed our recent ancestors are probably still in force today. So, to change the metaphor from a book to a cacophony of voices, the youngest voice, in its youthful vigour, might have something of a built-in advantage. Not an overriding advantage, however. I’d be content with the more cautious claim that the genetic book of the dead is a palimpsest made up of scripts ranging from very old to very young and including all intermediates between. If there are general rules governing relative prominence of old versus young or intermediate, they must wait for later research.
Biologists have long recognised morphological features that lie conservatively in basal layers of the palimpsest. An example is the vertebrate skeleton: the dorsally placed spinal column, with a skull and tail at the two ends, the column made of serially segmented vertebrae through which runs the body’s main trunk nerve. Then the four limbs that sprout from it, each consisting of a single, typically long bone (humerus or femur) connected to two parallel bones (radius/ulna, tibia/fibula); then a cluster of smaller bones terminating in five digits. It’s always five digits in the embryo, although in the adult some may be reduced or even missing. Horses have lost all but the middle digit, which bears the hoof (a massively enlarged version of our nail). A group of extinct South American herbivores, the Litopterns, included some species, such as Thoatherium (left), which independently evolved almost exactly the same hoofed limb as the horse (right). The two limbs have been drawn the same size for ease of comparison, but Thoatherium was considerably smaller than a typical horse, about the size of a small antelope. Think of the horse in the picture as a Shetland pony!
Litoptern
Horse
Arthropods have a different Bauplan (building plan or body plan), although they resemble vertebrates in their segmented pattern of units repeated fore-and-aft in series. Annelid worms such as earthworms, ragworms, and lugworms also have a segmented body plan, and they share with arthropods the ventral position of the main nerve. This difference in position of the body’s main nerve has led to the provocative speculation that we vertebrates may be descended from a worm who developed the habit of swimming upside down – a habit that has been rediscovered by brine shrimps today. If this is so, the ‘basic’ vertebrate Bauplan may not be quite as basic as we thought.
Brine shrimp
But, important and even stately as such morphological bauplans are, morphology has become overshadowed by molecular genetics when it comes to reading the lower layers of biological palimpsests in order to reconstruct animal pedigrees. Here’s a neat little example. South American trees are inhabited by two genera of tree sloths, the two-toed and the three-toed. There was also a giant ground sloth, which went extinct some ten or twelve thousand years ago, just recently enough to supply molecular biologists with DNA. Since the two tree sloths are so alike, in both anatomy and behaviour, it was natural to suppose that they are closely related, descended from a tree-dwelling ancestor quite recently, and more distantly related to the giant ground sloth. Molecular genetics now shows, however, that the two-toed tree sloth is closer to the giant sloth – all 4 tonnes of it – than it is to the three-toed tree sloth.
Long before modern molecular taxonomy burst onto the scene, morphological evidence aplenty showed us that dolphins are mammals not fish, for all that they look and behave superficially like large fish – mahi-mahi are indeed sometimes called ‘dolphinfish’ or even ‘dolphins’. But although science long knew that dolphins and whales were mammals, no zoologist was prepared for the bombshell released in the late twentieth century by molecular geneticists when they showed, beyond all doubt, that whales sprang from within the artiodactyls, the even-toed, cloven-hoofed ungulates. The closest living cousins of hippos are not pigs, as I was taught as a zoology undergraduate. They are whales. Whales don’t have hooves to cleave. Indeed, their land ancestors probably didn’t actually have cloven hooves, but broad four-toed feet, as hippos do today. Nevertheless, they are fully paid-up members of the artiodactyls. Not even outliers to the rest of the artiodactyls but buried deep within them, closer cousins to hippos than hippos are to pigs or to other animals who actually have cloven hooves. A staggering revelation that nobody saw coming. Molecular gene sequencing may have other shocks in store for us yet.
Just as a computer disc is littered with fragments of out-of-date documents, animal genomes are littered with genes that must once have done useful work but now are never read. They’re called pseudogenes – not a great name, but we’re stuck with it. They are also sometimes called ‘junk’ genes, but they aren’t ‘junk’ in the sense of being meaningless. They are full of meaning. If they were translated, the product would be a real protein. But they are not translated. The most striking example I know concerns the human sense of smell. It is notoriously poor compared with that of coursing hounds, seal-hunting polar bears, truffle-snuffling sows, or indeed the majority of mammals. You’d be right to credit our ancestors with feats of smell discrimination that would amaze us if we could go back and experience them. And the remarkable fact is that the necessary genes, large numbers of them, are still with us. It’s just that they are never read, never transcribed, never rendered into protein. They’ve become sidelined as pseudogenes. Such older scripts of the DNA palimpsest are not only there. They can be read in total clarity. But only by molecular biologists. They are ignored by the natural reading mechanisms of our cells. Our sense of smell is frustratingly poor compared to what it could be if only we could find a way to turn on those ancient genes that still lurk within us. Imagine the high-flown imagery that mutant wine connoisseurs might unleash. ‘Black cherry offset by new-mown hay in the attack, with notes of lead pencil in the satisfying finish’ would be tame by comparison.
Hippos are closer cousins to whales than to any other ungulates
The analogy between genome and computer disc is a more than usually close one. If I invite my computer to list the documents on my hard disc, I see an orderly array of letters, articles, chapters of books, spreadsheets of accounts, music, holiday photos, and so on. But if I were to read the raw data as it is actually laid out on the disc, I would face a phantasmagoria of disjointed fragments. What seems to be a coherent book chapter is made up of here a scrap, there a fragment, dotted around the disc. We think it’s coherent only because system software knows where to look for the next fragment. And when I delete a document, I may fondly imagine it has gone. It hasn’t. It’s still sitting where it was. Why waste valuable computer-time to expunge it? All that happens when you delete a document is that the system software marks its territory on the disc as available to be over-written by other stuff, as and when the space is needed. If the territory is not needed it will not be over-written and the original document, or parts of it, will survive – legible but never actually read – like the smell pseudogenes that we still possess but don’t use. This is why, if you want to remove incriminating documents from your computer, you must take special steps to expunge them completely. Routine ‘deletion’ is not proof against hackers.
Pseudogenes are a lucid message from the past: a significant part of the genetic book of the dead. If she hadn’t already deduced it from other cues, SOF would know, from the graveyard of dead genes littering the genome, that our ancestors inhabited a world of smells richer than we can imagine. The DNA tombstones are not only there, the lettering on them is more or less clear and distinct. Incidentally, these molecular tombstones are a huge embarrassment to creationists. Why on earth would a Creator clutter our genome with smell genes that are never used?
This chapter has been mainly concerned with deep layers of the palimpsest, the legacies of more ancient history. In the next four chapters we turn to layers nearer the surface. This amounts to a look at the power of natural selection to override the deep legacies of history. One way to study this is to pick out convergent resemblances between unrelated animals. Another way is ‘reverse engineering’. To which we now turn.
4 Reverse Engineering
One of the central messages of this book – that the meticulously detailed perfection we see in the external appearance of animals pervades the whole interior too – obviously rests on an assumption that something approaching perfection is there in the first place. There, and to be expected on Darwinian grounds. It’s an assumption that has been criticised and needs defending, which is the purpose of the next three chapters.
The most prominent critics of what they called ‘adaptationism’ were Richard Lewontin and Stephen Gould, both at Harvard, both distinguished, in their respective fields of genetics and palaeontology. Lewontin defined adaptationism as ‘That approach to evolutionary studies, which assumes without further proof that all aspects of the morphology, physiology and behavior of organisms are adaptive optimal solutions to problems.’ I suppose I am closer to being an adaptationist than many biologists. But I did devote a chapter of The Extended Phenotype to ‘Constraints on Perfection’. I distinguished six categories of constraint, of which I’ll mention five here.
Time lags (the animal is out of date, hasn’t yet caught up with a changing environment). Quadrupedal relics in the human skeleton supply one example.
Historical constraints that will never be corrected (e.g. recurrent laryngeal nerve, back-to-front retina).
Lack of available genetic variation (even if natural selection would favour pigs with wings, the necessary mutations never arose).
Constraints of costs and materials (even if pigs could use wings for certain purposes, and even if the necessary mutations were forthcoming, the benefits are outweighed by the cost of growing them).
Mistakes due to environmental unpredictability or malevolence (e.g. when a reed warbler feeds a baby cuckoo it is an imperfection from the point of view of the warbler, engineered by natural selection on cuckoos).
If such constraints are allowed for and admitted, I think I could fairly be called an adaptationist. There remains the point, which will occur to many people, that certain ‘aspects of the morphology, physiology and behavior of organisms’ may be too trivial for natural selection to notice them. They pass under the radar of natural selection. If we are talking about genes as molecular geneticists see them, then it is probably true that most mutations pass unnoticed by natural selection. This is because they are not translated into a changed protein, therefore nothing changes in the organism. They are literally neutral, in the sense of the Japanese geneticist Motoo Kimura, not mutations at all in the functional sense. It’s like changing the font in which an instruction is printed, from Times New Roman to Helvetica. The meaning is exactly the same after the mutation as it was before. But Lewontin had sensibly excluded such cases when he specified ‘morphology, physiology and behavior’. If a mutation affects the morphology, physiology, or behaviour of an animal, it is not neutral in the trivial ‘changing the font’ sense.
Nevertheless, some people still have an intuitive feeling that many mutations are probably still negligible, even if they really do affect morphology, physiology, or behaviour. Even if there’s a real change visible in the animal’s body, mightn’t it be too trivial for natural selection to bother about? My father used to try to persuade me that the shapes of leaves, say the difference between oak shape and beech shape, couldn’t possibly make any difference. I’m not so sure, and this is where I tend to part company with the sceptics like Lewontin. In 1964, Arthur Cain (my sometime tutor at Oxford) wrote a polemical paper in which he forcefully (some might say too forcefully) argued the case for what he called ‘The Perfection of Animals’. On ‘trivial’ characters, he argued that what seems trivial to us may simply reflect our ignorance. ‘An animal is the way it is because it needs to be’ was his slogan, and he applied it both to so-called trivial characters and to the opposite – fundamental features like the fact that vertebrates have four limbs and insects have six. I think he was on firmer ground where so-called trivial characters were concerned, for instance in the following memorable passage:
But perhaps the most remarkable functional interpretation of a ‘trivial’ character is given by Manton’s work on the diplopod [a kind of millipede] Polyxenus, in which she has shown that a character formerly described as an ‘ornament’ (and what could sound more useless?) is almost literally the pivot of the animal’s life.
Even in those cases where the character is very close to being genuinely trivial, natural selection may be a more stringent judge than the human eye. What is trivial to our eyes may still be noticed by natural selection when, in Darwin’s words, ‘the hand of time has marked the long lapse of ages’. JBS Haldane made a relevant hypothetical calculation. He assumed a selection pressure in favour of a new mutation so weak as to seem trivial: for every 1,000 individuals with the mutation who survive, 999 individuals without the mutation will survive. That selection pressure is much too weak to be detected by scientists working in the field. Given Haldane’s assumption, how long will it take for such a new mutation to spread through half the population? His answer was a mere 11,739 generations if the gene is dominant, 321,444 generations if it is recessive. In the case of many animals, that number of generations is an eye-blink by geological standards. A relevant point is that, however seemingly trivial a change may be, the mutated gene has very many opportunities to make a difference – via all the thousands of individuals in whose bodies it finds itself over geological time. Moreover, even though a gene may have only one proximal effect, because embryology is complicated, that one primary effect may ramify. As a result, the gene appears to have many seemingly disconnected effects in different parts of the body. These different effects are called pleiotropic, and the phenomenon is pleiotropism. Even if one of a mutation’s effects was truly negligible, it’s unlikely that all its pleiotropic effects would be.
With all due recognition to the various constraints on perfection, I think a fair working hypothesis is one that, surprisingly, Lewontin himself expressed, admittedly long before his attacks on adaptationism: ‘That is the one point, which I think all evolutionists are agreed upon, that it is virtually impossible to do a better job than an organism is doing in its own environment.’
Some biologists prefer to say natural selection produces animals that are just ‘good enough’ rather than optimal. They borrow from economists the term ‘satisficing’, a jargon word that they love to namedrop. I’m not a fan. Competition is so fierce, any animal who merely satisficed would soon be out-competed by a rival individual who went one better than satisficing. Now, however, we have to borrow from engineers the important notion of local optima. If we think of a landscape of perfection where improvement is represented by climbing hills, natural selection will tend to trap animals on the top of the nearest relatively low hill, which is separated from a high mountain of perfection by an impassable valley. Going down into the valley is the metaphor for getting temporarily worse before you can get better. There are various ways, known to both biologists and engineers, whereby hill-climbers can escape local optima and make their way to ‘broad, sunlit uplands’, though not necessarily to the highest peak of all. But I shall leave the topic now.
Engineers assume that a mechanism designed by somebody for a purpose will betray that purpose by its nature. We can then ‘reverse engineer’ it to discern the purpose that the designer had in mind.
Reverse engineering is the method by which scientific archaeologists reconstructed the purpose of the Antikythera mechanism, a mesh of cogwheels found in a sunken Greek ship dating from about 80 BC. The intricate gearing was exposed by modern techniques such as X-ray tomography. Its original purpose has been reverse engineered as an ancient equivalent of an analogue computer, designed to simulate the movement of heavenly bodies according to the system of epicycles later associated with Ptolemy.
Reverse engineering assumes that the object facing us had a purpose in the mind of a competent designer, a purpose that can be guessed. The reverse engineer sets up a hypothesis as to what a sensible designer might have had in mind, then checks the mechanism to see if it fits the hypothesis. Reverse engineering works well for animal bodies as well as for man-made machines. The fact that the latter were deliberately designed by conscious engineers while the former were designed by unconscious natural selection makes surprisingly little difference: a potential for confusion readily exploited by creationists with their characteristically eager appetite for it. The grace of a tiger and of its prey could not easily, it would seem, be bettered:
What immortal hand or eye Could frame thy fearful symmetry.
Indeed, animals sometimes seem too symmetrically designed, to their own detriment: remember the owl pictured in Chapter 2.
Darwin had a section of Origin of Species called ‘Organs of extreme perfection and complication’. It’s my belief that such organs are the end products of evolutionary arms races. The term ‘armament race’ was introduced to the evolution literature by the zoologist Hugh Cott in his book on Animal Coloration published in 1940, during the Second World War. As a former officer in the regular army during the First World War, he was well placed to notice the analogy with evolutionary arms races. In 1979, John Krebs and I revived the idea of the evolutionary arms race in a presentation to the Royal Society. Whereas an individual predator and its prey run a race in real time, arm races are run in evolutionary time, between lineages of organisms. Each improvement on one side calls forth a counter-improvement on the other. And so the arms race escalates, until called to a halt, perhaps by overwhelming economic costs, just like military arms races.
Antelopes could always outrun lions, and vice versa, but only by counter-productive investment of too much ‘capital’ in leg muscles at the expense of other calls on investment in, say, milk production. If the language of ‘investment’ sounds too anthropomorphic, let me translate. Individuals who excel in running speed would be out-competed by slightly slower individuals who divert resources more usefully, from athletic legs into milk. Conversely, individuals who overdo milk production are out-competed by rivals who economise on milk production and put the energy saved into running speed. To quote the economists’ hackneyed saw, there’s no such thing as a free lunch. Trade-offs are ubiquitous in evolution.
I think arms races are responsible for every biological design impressive enough to, in the words of David Hume’s Cleanthes, ravish ‘into admiration all men who have ever contemplated them’. Adaptations to ice ages or droughts, adaptations to climate change, are relatively simple, less prone to ravish into admiration because climate is not out to get you. Predators are. So are prey, in the indirect sense that, the more success prey achieve at evading capture, the closer their would-be predators come to starvation. Climate doesn’t menacingly change in response to biological evolution. Predators and prey do. So do parasites and hosts. It is the mutual escalation of arms races that drives evolution to Cleanthean heights, such as the feats of mimetic camouflage we met in Chapter 2, or the sinister wiles of cuckoos that will amaze us in Chapter 10.
And now for a point that at first sight seems negative. Whereas animals look beautifully designed on the outside, as soon as we cut them open, we seem superficially to get a different impression. An untutored spectator of a mammal dissection might fancy it a mess. Intestines, blood vessels, mesenteries, nerves seem to spill out all over the place. An apparent contrast with the sinewy elegance of, say, a leopard or antelope when seen from outside. On the face of it, this might seem to contradict the conclusion of Chapter 2. The central point stated there was that the perfection typical of the outer layer must pervade every internal detail as well. Now compare your heart with the village pump, which seems neatly and simply fit for purpose. Admittedly, the heart is two pumps in one, serving the lungs on the one hand and the rest of the body on the other. But you could be forgiven for wondering whether a more minimally elegant pump might profitably have been designed.
Each eye sends information to the brain on the opposite side. Muscles on the left side of the body are controlled by the right side of the brain and vice versa. Why? I suppose we are again dealing with ancient scripts long buried in low strata of the palimpsest. Given such deep constraints, natural selection busily tinkers with the upper-level scripts, making good, as far as possible, the inevitable imperfections imposed by deeper levels. The backwards wiring of the vertebrate retina is well compensated by post-hoc making good. You might think that ‘from such warped beginnings nothing debonair can come’. The great German scientist Hermann von Helmholtz is said to have remarked that if an engineer had produced the eye for him, he would have sent it back. Yet after tweaking, ‘in post’ as movie-makers say, the vertebrate eye can become a fine piece of optical kit.
Two pumps
Why do animals look obviously well designed on the visible outside but apparently less so inside? Does the clue reside in that word ‘visible’? In the case of Chapter 2’s camouflage, and also ornamental extravaganzas like the peacock’s fan, (human) eyes are admiring the external appearance of the animal, and (peahen or predator) eyes are doing the natural selection of external appearance: similar vertebrate eyes in both cases. No wonder external appearance looks more perfectly ‘designed’ than internal details. Internal details are every bit as subject to natural selection, but they don’t obviously look that way because it is not selection by eyes.
That explanation won’t do for the streamlined flair of a sprinting cheetah, or its equally graceful Tommy prey. Those beauties did not evolve for the delectation of eyes but to satisfy the lifesaving requirements of speed. Here it would seem to be the laws of physics that impose what we perceive as elegance: as it is for the aerodynamic grace of a fast jet plane. Aesthetics and functionality converge on the same stylish elegance.
I confess that I find the interior of the body bewilderingly complex. I might even go so heretically far as to dismiss it as a mess. But I am a naive amateur where internal anatomy is concerned. A consultant surgeon whom I have consulted (what else should one do with a consultant?) assures me in no uncertain terms that, to his trained eye, internal anatomy has a beautiful elegance, everything neatly stowed away in its proper place, all shipshape and Bristol fashion. And I suspect that ‘trained eye’ is exactly the point. In Chapter 1, I contrasted the ear’s effortless deciphering of the spoken word ‘sisters’ with the eye’s fumbling impotence to see anything beyond a wavy line on an oscilloscope. My eye sees elegance on the outside. Then when I cut an animal open, my amateur eye contemplates only a mess. The trained surgeon sees stylish perfection of design, inside as well as out. It is, at least partly, the story of ‘sisters’ all over again. Yet there is more to be said. Something about embryology.
Veins, nerves, arteries, lymphatic system – a whole armful of complexity
The sceptic vocally doubts whether it can really matter whether this vein in the arm passes over or under that nerve. Maybe it doesn’t in the sense that, if their relationship could be reversed with a magic wand, the person’s life might not suffer, and might even improve. But I think it does matter in another sense – the sense that solved the riddle of the laryngeal nerve. Every nerve, blood vessel, ligament, and bone got that way because of processes of embryology during the development of the individual. Exactly which passes over or under what may or may not make a difference to their efficient working, once their final routing is achieved. But the embryological upheaval necessary to effect a change, I conjecture, would raise problems, or costs, sufficient to outweigh other considerations. Especially if the embryological upheaval strikes early. The intricate origami of embryonic tissue-folding and invagination follows a strict sequence, each stage triggering its successor. Who can say what catastrophic downstream consequences might flow from a change in the sequence – the kind of change necessary to re-route a blood vessel, say.
Moreover, perhaps Darwinian forces have worked on human perception to sharpen our appreciation of external appearances as opposed to internal details. At all events, I revert with confidence to the conclusion of Chapter 2. It is entirely unreasonable to suppose that the chisels of natural selection, so delicately adept at perfecting external and visible appearance, should suddenly stop at the animal’s skin rather than working their artistry inside. The same standards of perfection must pervade the interior of living bodies, even if less obviously to our eyes. To dissect the non-obvious and make it plain will be the business of future zoological reverse engineers, and it is to them that I appeal.
Ideally, reverse engineering is a systematic scientific project, perhaps involving mathematical models in the sense discussed in Chapter 1. More usually, at present at least, it involves intuitive plausibility arguments. If the object in question has a lens in front of a dark chamber, focusing a sharp image on a matrix of light-sensitive units at the back of the chamber, any person living after the invention of the camera can instantly divine the purpose for which it evolved. But there will be numerous details that will matter and will require sophisticated techniques of reverse engineering, including mathematical analysis. In this chapter our reverse engineering is mostly of the intuitive, common sense kind, like the example of the eye and the camera.
Reverse engineering is supplemented by comparison across species. If SOF is confronted with a hitherto unknown animal, she can read it both by pure reverse engineering (‘a device designed by an engineer to do such-and-such would probably look rather like this’) and also by comparison with known species (‘this organ looks like an organ in so-and-so species that we already know, and it probably is used for the same purpose’).
An indirect version of reverse engineering can be used to infer aspects of an animal that cannot be seen, for example when all we have is fossils. We have no fossil evidence about the heart of a dinosaur. But fossils tell us that some sauropods such as Brontosaurus and the even larger Sauroposeidon had extraordinarily long necks. The CGI artists of Jurassic Park beautifully illustrated the dominant view that they reached up to browse tall trees. Like giraffes, only more so. Now the engineer steps in and invokes simple laws of physics to dictate that the heart would have had to generate very high pressure in order to push blood to the height of the animal’s brain when plucking leaves from a high tree. You can’t suck water through a straw that’s more than 10.3 metres tall, even if your sucking is powerful enough to generate a perfect vacuum in the straw. Sauroposeidon’s head probably overtopped its heart by about that much, which gives an idea of the pressure that the heart would have had to generate to push blood up to the head. Without ever seeing a fossilised sauropod heart, the engineer infers that it must have generated especially high pressure. Either that or that they didn’t browse trees at all.
I can’t resist reflecting that the difficulty of pumping blood to a head so high might have been partially responsible for those large dinosaurs outsourcing some brain functions to a second ‘brain’, in the pelvis. Also, I never miss an excuse to quote Bert Leston Taylor’s delightfully witty poem on the subject.
Behold the mighty dinosaur, Famous in prehistoric lore, Not only for his power and strength But for his intellectual length.
You will observe by these remains
The creature had two sets of brains –
One in his head (the usual place),
The other at his spinal base,
Thus he could reason A priori
As well as A posteriori.
No problem bothered him a bit
He made both head and tail of it.
So wise was he, so wise and solemn,
Each thought filled just a spinal column.
If one brain found the pressure strong
It passed a few ideas along.
If something slipped his forward mind
’Twas rescued by the one behind.
And if in error he was caught
He had a saving afterthought.
As he thought twice before he spoke
He had no judgment to revoke.
Thus he could think without congestion
Upon both sides of every question.
Oh, gaze upon this model beast,
Defunct ten million years at least.
The pelvic ‘brain’ would have been about on a level with the heart, and impressively much lower than the head.
Alas, there are no sauropods for us to test such ideas, and we must make do with the next best thing, which is the giraffe. Though not in the same league as a giant dinosaur, the giraffe’s head is quite lofty enough to require an abnormally high blood pressure, out of the ordinary for a mammal. And the following graph bears out the expectation.
I have plotted mean arterial blood pressure against the logarithm of body mass for a range of mammals from mouse to elephant. It’s best to use logarithms for the weights – otherwise it would be hard to fit mouse and elephant on the same page, with intermediate animals conveniently spread out between. The dotted line is the straight line that best fits the data. The line slopes upwards – larger animals tend to have higher blood pressure. Most species are pretty close to the line, meaning that their blood pressure is close to typical for their weight. But the big exception is the giraffe, which is far above the line. Its blood pressure is way higher than it ‘should be’ for an animal of its size. Surprisingly, other evidence shows that the giraffe heart is not especially large. It seems to be prevented from enlarging in evolution by the need to share the body cavity with large herbivorous guts. It achieves the extra-high blood pressure in a different way, by a greater density of heart muscle cells, an improvement that probably imposes costs of its own. Without ever seeing a Brontosaurus heart, we can predict that it too would have stood way above the line in the equivalent graph for reptiles.
The teeth of a hitherto unknown animal speak volumes, and this is fortunate because teeth, being necessarily hard enough to crunch food, are also hard enough to outlast anything else in the fossil record. Some important extinct species are known only from teeth. In the rest of this chapter, we shall use teeth and other biological food-processing devices as our example of choice. Look at this ancient skull. The first thing you notice is the scary canine teeth. You might reverse engineer these as being good for either fighting rivals or stabbing prey to death and holding onto them. Seeking further evidence, you might then look at the other teeth near the back of the jaw, the molars. They don’t mesh surface-to-surface in the way that ours or a horse’s do, but shear past each other like scissors as the jaws close. They seem designed to slice rather than to mill. This says ‘carnivore’. Well, obviously. But it’s only obvious because we are rather good at intuitive reverse engineering, and because we have living large carnivores like lions and tigers for comparison. It does no harm to make the reasoning explicit.
Sabretooth
Animals, perhaps because they are themselves made of meat, find meat relatively easy to digest, and carnivore intestines tend to be appropriately short. If SOF were handed an unknown animal, very long intestines would signal ‘herbivore’ to her. I’ll return to this. Meat, moreover, demands relatively little pre-processing with teeth before digestion. Cutting off substantial chunks to be swallowed whole is sufficient. Plants may be easier to catch than animals – they don’t run away – but they make up for it by being harder to process once you’ve caught them. Plant cells are different from animal cells. They have thick walls toughened by cellulose and silica. For this and other reasons, herbivores need to grind their food into tiny pieces before it is ready to pass into the gut for further breaking up chemically into even smaller pieces. Herbivore teeth are millstones which, like the mills of God, grind slowly and they grind exceeding small. Carnivore teeth don’t resemble millstones and they don’t grind. They cut, shearing through fibrous tissues.
Looking at the back teeth of the above skull, then, we confirm our initial diagnosis from the dagger-like canines, and convincingly reverse-engineer our scary specimen as telling a tale of ancestral carnivores. Moving to the rest of the skull, we note that the articulation of the lower jaw allows only up-and-down movement suitable for scissoring food, not side-to-side movement such as would be needed for milling. Up and down is putting it mildly: the sheer size of the gape is formidable. As you’ll have guessed, this is the skull of a sabretooth cat, often called sabretooth tiger, although it could just as well be called sabretooth lion. It was a big cat, Smilodon, not closer to any particular modern big cat than to any other. Contemporaneous with Smilodon, there were true lions in America, now extinct, bigger than Smilodon, bigger than African lions.
How did Smilodon use those formidable fangs? It’s notable that among modern carnivores, the cat family (Felidae) runs to long canine teeth more than the dog family (Canidae), despite the name ‘canine’ for the teeth. A plausible reason is as follows. Canids are mostly pursuit-hunters. They run their prey down to exhaustion. When they finally catch up with it, the poor spent creature is in no state to escape. Killing it is not a problem. Just start eating! Felids, on the other hand, tend to be stalkers and ambushers. Their prey, when they first pounce upon it, is fresh and in a strong position to escape. Either a swift killing stab or an inescapable grip is desirable, and long penetrating canines answer both needs. Among living cats, the clouded leopard sports the nearest approach to the sabres of Smilodon. Clouded leopards spend much of their time in trees and drop on their prey. Long, sharp daggers would be especially suited to subduing an animal taken by surprise from above, not ‘heated in the chase’ and in full possession of its powers.
Turning to other parts of the skull of Smilodon, we notice that the eye sockets point forward, indicating binocular vision, useful for pouncing on prey and no good for seeing danger creeping up from behind. Sabretooths had no need to watch their back. Herbivorous animals, whose ancestors became ancestors by virtue of noticing would-be killers, tend to have lookout eyes pointing sideways, giving almost 360° vision, calculated to spot a predator stalking from any direction.
Clouded leopard
So now, suppose you are presented with the skull below. It’s obviously very different. The eyes look sideways, as if scanning all around for danger while not being especially concerned with what is ahead. Probably an animal with a need to fear predation, then. The incisor teeth at the front look well suited to cropping grass. Most noticeable are the back teeth. They are broad grinders rather than sharp slicers, and they meet their opposite numbers in a precise fit when the jaws close. Their whole shape with its articulation is well suited to grinding plant food into very small pieces, again confirming the suspicion that this animal’s genes survived in a world of grass or other plant food. And the lower jaw, unlike that of Smilodon, moves sideways as well as up and down, a good milling action. This fossil is Pliohippus, an extinct horse that lived in the Pliocene, probably in mortal fear of Smilodon.
Pliohippus
The contrast between the skulls of the carnivorous sabretooth and the herbivorous horse is stark and clear. There was an animal called Tiarajudens, one of those we used to call a mammal-like reptile (nowadays we’d call it an early mammal), which flourished perhaps 280 million years ago, before the great age of dinosaurs. It had impressive sabretooth canines, much like Smilodon, which indicate a carnivorous diet similar to that of the formidable cat. But the back teeth suggest that, along with other animals to whom it was related, it was in fact a herbivore. So, we have a mismatch. Why would a creature with grinding back teeth have canine teeth like Smilodon? Perhaps Tiarajudens was a herbivore equipped with daggers for defence against predators. Or perhaps, like modern walruses, for fighting against rivals of its own species, as elephants use their gigantic tusks (elephant tusks are enlarged incisor teeth, not canines as in walruses).
Walrus
Walruses have been seen using their (upper canine) tusks to lever themselves out of the water and to make holes in the ice. Anyway, Tiarajudens stands as a cautionary warning against over-hasty reverse engineering, looking at only one thing, in this case the canine teeth.
Hedgehog
Some mammals such as shrews and small bats eat insects. Dolphins eat fish. Though technically carnivorous, the dental demands of these diets are different. Insectivorous teeth are neither grinders nor cutters but piercers. They tend to have sharp points, well suited to piercing the external skeletons of insects. If SOF’s unknown specimen sported piercing teeth like those of this hedgehog, she’d suspect that its ancestors survived on a diet of insects and other arthropods. And that is correct, but they like earthworms too. Ants and termites are a special case (see below).
Common dolphin
Gavial
And now here’s the skull of a dolphin (top), and a gavial (bottom), to show typical fish-eating teeth and jaws. These two fish-eaters, a mammal and a crocodilian, have independently evolved pretty much the same dentition and jaw shape, an example of convergent evolution (which is the topic of Chapter 5). What’s the reverse-engineering explanation for this convergent resemblance? Fish-eaters, unlike, say, lions, are usually much larger than their prey. They don’t need to grind or cut or pierce their prey. Their prey is small enough to swallow whole. Long rows of small, pointed teeth are well equipped to grasp a slippery, soft fish and prevent it from escaping. And the slender jaws can snap shut on the fish without expelling a rush of water that might propel it out of harm’s way.
Ichthyosaur
If you were lucky enough to stumble upon a fossil like the above, you could apply the lesson of the previous paragraph: fish-eater. It’s an ichthyosaur such as we met in Chapter 3, a contemporary and relative of dinosaurs, member of a large group that went extinct somewhat earlier than the last of the dinosaurs. Both reverse engineering, and comparison with the dolphin and gavial pictures, speak to us loud and clear: its ancestors ate fish.
Killer whales (Orca) and sperm whales can be thought of as giant dolphins. They too eat prey smaller than themselves, and they too have long rows of dolphin-like teeth but hugely enlarged. Sperm whales have them only in the lower jaw (very occasionally in the upper jaw, and we may take this as a vestigial relic). Killer whales have them in both jaws. All other large whales, the so-called baleen whales, are filter feeders, sieving krill (crustaceans). They have no teeth at all (though, revealingly, their embryos have them and never use them). Their huge baleen filters are made of keratin, like hooves, fingernails, and rhinoceros horn. The reverse engineer would have no trouble in diagnosing a baleen whale as a trawler. Actually, they are better than trawlers, for they will target a huge aggregation of krill, and gulp it in with copious quantities of sea water, which is then forced out through the curtain of baleen, trapping the krill.
Ants and termites are colossally numerous. A specialist capable of penetrating an ant nest’s formidable defences can hoover up a bonanza of food denied to an ordinary insectivore like a hedgehog. And their dentition is correspondingly specialised. For this purpose, by the way, termites are honorary ants. Mammals who preferentially eat ants and/or termites are all called anteaters. There’s a group of three South American mammals whose name in English is ‘anteater’: the Giant Anteater, the Lesser Anteater, and the Silky Anteater.
Giant Anteater
Tamandua
Giant Anteater
Pangolin
Armadillo
Echidna
The Giant Anteater’s scientific name, Myrmecophaga, is simply Greek for ‘anteater’. You will already have concluded that, since other mammals also specialise in eating ants, ‘Anteater’ is not a great name for a taxonomic group. I’ll use a capital letter for the three South American ‘Anteaters’ and a lower-case letter for other mammals who eat ants (or termites).
The South American Anteaters push the anteating habit to its extreme. The skulls of two of them, Tamandua and the Giant Anteater Myrmecophaga, are pictured at the top of the page opposite. Notice the extreme prolongation of the snout and the total absence of teeth. You’d hardly recognise the Giant Anteater’s skull as a skull at all. All anteaters show the same features, if to a lesser extent. The pangolin has no teeth and a moderately long snout. Armadillos have a longer snout and rather small teeth. The aardvark or antbear of Africa has back teeth, but no teeth at all along most of its long snout. Myrmecobius, the numbat, marsupial anteater of Australia, has a long, pointy head. It has teeth but doesn’t use them for eating except in infancy. Adults seem to use them only for gripping and preparing nest material.
Tachyglossus, the spiny anteater or echidna of Australia and New Guinea, is as distant as you can get from all the above while still being a mammal. It’s an egg-laying mammal like the platypus, a leftover from the ‘mammal-like reptiles’ of the ancient supercontinent of Gondwana. But unlike the platypus, with which it shares deep palimpsest features, it does, as its English name suggests, eat ants and termites. And its rather weird-looking skull does indeed have a long, slender snout and no teeth. Let’s not get carried away, however. A slightly longer snout is possessed by the related echidna genus, Zaglossus, and Zaglossus eats almost nothing but earthworms. Evidently, we must be careful before we jump too precipitately to the conclusion that ‘long snout’ necessarily means anteater. Anteating is not the only habit capable of writing ‘long snout’ in the palimpsest.
What else might SOF use to diagnose an animal as an anteater? Myrmecophaga, the Giant Anteater of South America, whose hugely elongated skull we have already seen, has a giant-sized sticky tongue, which it can protrude to a length of 60 cm, having deployed its formidable claws to break into an ant or termite nest. Huge numbers of the insects stick to the tongue and are drawn in before the tongue shoots out again. Despite its great length, the tongue flicks out and in again at high speed, more than twice per second. Though none can quite match Myrmecophaga, creditably long, sticky tongues are also found, convergently evolved, in aardvarks and the unrelated aardwolves, who, unlike other members of the hyaena family, specialise in eating termites. Pangolins, too, have convergently evolved a long sticky tongue. That of the giant pangolin can be 40 cm long and is attached way back near the pelvis instead of to the hyoid bone in the throat, like ours. A pangolin can extend its tongue deep inside an ants’ nest, skilfully steering through the labyrinth of tunnels, turning left, turning right, leaving no subterranean avenue unexplored. Tamanduas also have a long sticky tongue but, in this case, their evolution was not independent of Myrmecophaga. They surely inherited the long tongue from their shared ancestor, also an anteater. The egg-laying spiny anteater too has a long, sticky tongue, and this time it really is convergent. As is that of the numbat, the marsupial anteater.
There are also physiological resemblances among anteating mammals, notably a low metabolic rate and low body temperature, convergently evolved enough times to impress our hypothetical SOF. However, a low metabolic rate is not exclusively diagnostic of an ant-eating habit. Sloths, befitting their name, also have a low metabolic rate. So do koalas, whom you could regard as a kind of marsupial equivalent of sloths. Both live up trees, eating relatively un-nutritious leaves, and both are slow moving, you might even say lethargic. The convergence doesn’t extend to both ends of the alimentary canal, however. Koalas defecate more than a hundred times per day, while sloths hold the record for the other extreme. They defecate about once per week, maybe because they laboriously climb down from the tree in order to do so.
Some of my reverse-engineering conjectures could be wrong. They are only provisional, to illustrate the point that the teeth of an animal, if properly read, will tell a story. In many cases, a story of ancient grassland prairies or leafy forests. Or, if the teeth resemble those of Smilodon or the clouded leopard, they speak to us of ambush and stalking. No doubt, if we could read them, every tooth we find could plunge us ever deeper into more specific, detailed stories. Teeth are enamelled archives of ancient history.
Teeth constitute the first food processor in the conveyor belt of digestion. The revealing differences between carnivores and herbivores continue on into the gut. Weight for weight, plants are not so nutritious as meat, so cows, for example, need to graze pretty continuously. Food passes through them like an ever-rolling stream, and they defecate some 40 or 50 kilograms per day. Plant stuff being so different from their own bodies, herbivores need help from chemical specialists to digest it. Those specialist chemists, some of whom were honing their skills perhaps a billion years before animals came on the scene at all, include bacteria, archaea (formerly classified as bacteria but actually far separated from them), fungi, and (what we used to call) protozoa. Ruminants such as cows and antelopes do their fermentation in a different way from horses and rabbits, and at different ends of the gut, but all rely on help from micro-organisms. As already mentioned above, herbivores have longer guts than carnivores, and their guts are complicated by elaborate blind alleys and fermentation chambers, specially fashioned to house symbiotic micro-organisms. Ruminants have the added complication of sending the food back for reprocessing by the teeth for a second time after it’s been swallowed – chewing the cud.
Herbivore gut
Carnivore gut
There is one bird, the hoatzin of South America, which eats nothing but leaves, the only bird to do so. And – an example of convergent evolution, the process we’ll meet in the next chapter – the hoatzin resembles ruminant mammals in having lots of little gut chambers in which are housed bacteria wielding the necessary chemical expertise to digest leaves. Incidentally, there’s a widely believed myth that the hoatzin is unique among birds in retaining ancient claws in the front of the wing, like the Jurassic ‘intermediate’ fossil Archaeopteryx. It’s true that hoatzin chicks have these primitive claws, but so do the chicks of many other birds, as David Haig pointed out to me. He went on to suggest that this mythic meme is popular among both biologists and creationists, who respectively want Archaeopteryx to be, and not to be, an ‘evolutionary intermediate’. No animal exists to be primitive for the sake of it, nor to serve as an evolutionary intermediate. The claws are useful to the chicks, who used them for clambering back into a tree when they fall.
Tiktaalik
By the same token animals don’t exist for the sake of ‘moving on to the next stage in evolution’. The Devonian fossil Tiktaalik is widely touted as a transition between fish and land vertebrates. So it may be, but being transitional is not a way to earn a living. Tiktaalik was a living, breathing, feeding, reproducing creature, which should be reverse-engineered as such – not as a half-way stage on the way to something better.
What of our own teeth and jaws, our own guts, and those of our near relatives? What tales of long-gone ancestral meals do they tell? Comparison of our Homo sapiens lineage with extinct hominins such as Paranthropus (Australopithecus) robustus and boisei shows a marked trend over time towards shrinkage of both jaws and teeth in our sapiens lineage. The ribcage of those robust old hominins could accommodate a large vegetarian gut. They were evidently less carnivorous than we are, equipped with large plant-milling teeth, strong grinding jaws, and correspondingly powerful jaw muscles. Even though the muscles themselves have not fossilised, their bony attachments, sometimes culminating in a vertical (‘sagittal’) crest like a gorilla’s to increase their purchase, speak to us eloquently of generations of plant roughage. Our own jaw muscles don’t reach so high up the side of our head and we have no bony crest.
The primatologist Richard Wrangham has promoted the intriguing hypothesis that the invention of cooking was the key to human uniqueness and human success. He makes a persuasive case that our reduced jaws, teeth, and guts are ill-suited to either a carnivorous or a herbivorous diet unless a substantial proportion of our food is cooked. Cooking enables us to get energy from foods more quickly and efficiently. For Wrangham it was cooking that led to the dramatic evolutionary enlargement of the human brain, the brain being by far the most energy-hungry of our organs. If he’s right, it’s a nice example of how a cultural change (the taming of fire) can have evolutionary consequences (the shrinking of jaws and teeth).
Birds have no teeth, nor bony jaws. Surprising as it sounds, they may have lost them to save weight – an important concern in a flying animal – replacing them with light, horny beaks. The word ‘mandible’ is used for both parts of the beak – the upper mandible and the lower mandible. Beaks can tear but they can’t chew. Birds do the equivalent of chewing with the gizzard, a muscular chamber of the gut, often containing hard gastroliths – stones or grains of sand that the bird swallows to help with the milling process. Ostriches swallow appropriately large stones, up to 10 cm. Being flightless, they don’t have to worry so much about weight. Even larger stones found with fossil birds such as the giant moas of New Zealand are identified as gastroliths by their polished surfaces – polished by the grinding action in the gizzard.
1. Macaw
2. Crossbill
3. Spoonbill
4. Eagle
5. Skimmer
6. Hummingbird
Beaks vary greatly, and speak to us eloquently of different ways of procuring food. Their variety has been compared with the set of pliers in a mechanic’s toolkit. Pointed beaks delicately select small targets such as single seeds or grubs. Parrot beaks are robust nutcrackers or large seed crushers, and the curved upper mandible with its pointed tip is used as something like a hand. Caged parrots can often be seen climbing on the bars, levering themselves up with the beak as if it were a hand. In the wild they use the same trick in trees. Hummingbird beaks are long tubes for imbibing nectar. Imperious, hooked eagle beaks rip flesh from carcases. Woodpecker beaks hammer like high-powered pneumatic drills, pounding rhythmically into trees in search of larvae. They have specially reinforced skulls to cope with the shock of hammering. Flamingo beaks are upside-down filters for small crustaceans, the bird world’s nearest approach to the krill-sieving baleen of whales. Oystercatchers use their long, pointed beaks to chisel into mussels and other shellfish. Curlews use theirs to probe mud for worms and shellfish. Spoonbills have flat paddle-like bills that they sweep from side to side, at the same time using their feet to stir up mud and expose small animals lurking in it. Skimmer beaks are even more specialised. The lower mandible is longer than the upper. The bird flies close to the water with the mouth open and the tip of the lower mandible skimming the surface. When it hits a fish, the beak snaps shut, trapping the fish. Pelicans have a voluminous pouch of skin under the beak, which nets fish.
Nestling birds who are fed by their parents don’t need beaks to do anything other than gape. Their beaks are grotesquely wide, with brightly coloured linings – advertising surfaces garishly designed to out-compete their siblings for parental largesse. The huge difference from adult beaks of the same species reminds us that juvenile needs can be very different from adult ones, a principle writ large by caterpillars and butterflies, tadpoles and frogs, and many other examples where larval forms occupy a completely different niche from the adults they become.
GALAPAGOS FINCHES
Large ground finch
Medium ground finch
Small tree finch
Green warbler finch
Crossbills sport a weird crossover of upper and lower jaw beaks, which is helpful in prying apart the scales of pinecones. Insectivorous birds have differently shaped beaks from seed-eaters. And specialists on seeds of different sizes have correspondingly different beaks, the differences making total sense from a reverse-engineering point of view. The evolution of such differences is the subject of a beautiful and still proceeding long-term study of ‘Darwin’s Finches’ on one of the smaller Galapagos Islands by Peter and Rosemary Grant, and their collaborators.
Galapagos is matched as a Pacific island showcase of Darwinian evolution by the archipelago of Hawaii. Both island chains are volcanic and very young by geological standards. The biology of Hawaii differs in being more contaminated by humans, and by the other invasions for which humans are to blame. The evolutionary divergence of Hawaiian honeycreepers (below) shows a variety of beaks that outdoes even that of the Galapagos finches (above). There are eighteen surviving species (more than twice that number have gone extinct), all apparently descended from a single species of Asian finch, probably looking not unlike a Galapagos finch. The range of bill types that has evolved in such a short time is astonishing.
Some have retained the seed-eating habits of the ancestor, and still look finch-like with stout, stubby beaks. Others have modified their beaks for nectar-sipping, like African sunbirds rather than like New World hummingbirds. Yet others, with long downward-curving beaks, are probers for insects. Of these, the so-called ‘I’iwi’ (below) has a sharp, stout, stabbing lower mandible, which hammers into bark. Then the long curved upper mandible, which has been held out of the way during the hammering, comes into action to probe insects out of the cracks. The Maui parrotbill uses its powerful callipers to crush twigs and rip off bark in search of insects.
HAWAIIAN HONEYCREEPERS
Laysan finch
Kakawahie (extinct)
‘Akiapola’au
‘I’iwi
Heron beaks are long fishing spears, stabbing down into the water with sudden precision. The African black heron uses its wings to shade its field of view, which would otherwise be troubled by reflections from the rippling water surface. It dramatically sweeps its black wings across its body, laughably recalling a black-cloaked villain in Victorian melodrama. A separate problem for anyone spearfishing from above is refraction at the water surface – the illusion that makes oars look bent. There is some evidence that herons and kingfishers adjust their aim to compensate. The archer fishes of Southeast Asia face the same problem in reverse. They lurk under water and shoot insects sitting on tree branches above the surface, by squirting a sudden jet of water straight at the target. That’s remarkable enough in itself. Even more so, they seem to compensate for refraction, like herons but in the other direction.
Archer fish
Reverse engineering, then, is one method by which we can read the body of an animal. Another method is to compare it with other animals, both related and unrelated. We used this method to some extent in this chapter. When the genetic books of unrelated animals spell the same message about their environment and way of life, we call it convergence. Convergent resemblances can be spectacular, as we’ll see in the next chapter.
5 Common Problem, Common Solution
This book’s main thesis is that every animal is a written description of ancestral worlds. It rests upon the hidden assumption – well, not so very hidden – that natural selection is an immensely powerful force, carving the gene pool into shape, deep down among the smallest details. As we saw in Chapter 2, among the most convincing evidence for the power of natural selection is the perfection of camouflage, the consummate detail with which some animals resemble their (ancestral) environment, or resemble an object in that environment. Equally impressive is the detailed resemblance of an animal to another, unrelated animal, because both have converged on the same way of life. Matt Ridley’s How Innovation Works documents how our greatest human innovations have been hit upon many times independently by inventors in different countries, working in ignorance of each other’s efforts. Just the same is true of evolution by natural selection. This chapter is about convergent evolution as an eloquent witness to the power of natural selection.
Despite appearances, the animal above opposite is not a dog. It is an unrelated marsupial, Thylacinus, the Tasmanian wolf (often called Tasmanian tiger, for no better reason than the stripes). In (what hindsight can now see as) a heinous crime against nature, the Tasmanian government in 1888 put a bounty on thylacine heads. The last one to be seen in the wild was infamously shot in 1930 by someone called Wilf Batty. He must have known it was almost extinct, though he couldn’t have known his victim was the last one. I suppose in 1930 people still didn’t care about such things, a poignant example of what I have called the shifting moral Zeitgeist. A captive specimen called Benjamin survived in Hobart Zoo until 1936. Thylacinus is one of the best-known examples of convergence. It looked like a dog because it had the same way of life as a dog. Its skull especially is so like a dog’s that it is a favourite trick question in zoology student examinations. Such a favourite, indeed, that in my year at Oxford they gave us a real dog skull as a double bluff, assuming that we’d automatically plump for Thylacinus.
Thylacine
Rhinoceros beetle
You’d never mistake this for a rhinoceros. But if you watched two rhinoceros beetles fighting, and then two rhinoceroses, you’d realise that convergent resemblances can vault over many orders of magnitude of body size. A fight is a fight is a fight, and a horn is a handy weapon at any size. The same goes for stag beetles and stags, with a somewhat dramatic embellishment. Stag beetles, but not stags, can lift their rivals high in the air on the prongs of their ‘antlers’.
Paca
Chevrotain
On the left is a paca, a rodent from the rainforests of South and Central America. To its right is a chevrotain or ‘mouse deer’, an even-toed ungulate that lives in Old World forests. They look like each other convergently because they have similar ways of life. In Africa, the niche is filled by a small ungulate, in South America, by a large rodent.
Armadillos are South American mammals, armoured against predators. When threatened, they roll up into a ball. The picture to the left shows the three-banded armadillo, which rolls up with especially compact elegance. In one of its illustrative quotations, the Oxford English Dictionary startlingly records that ‘Formerly the armadillo was used in medicine, being swallowed as a pill in its rolled-up state.’ Quite a stretch! Until you realise that ‘armadillo’ in this 1859 quotation referred not to the mammal but to a convergent crustacean, a woodlouse, whose Latin name Armadillidium means ‘little armadillo’. Armadillo itself is a Spanish word, a diminutive of armado or ‘armed’. So Armadillidium is a diminutive of a diminutive, a double diminutive. The commonality of name speaks to the power of convergent evolution. As befits its vernacular name of ‘pill bug’, in its rolled-up state you could indeed swallow a woodlouse whole, although as to its alleged medicinal value, I shall not comment. The mammalian armadillo and the crustacean Armadillidium have converged in their evolution, independently hitting on the same protective habit, albeit at very different sizes, rolling themselves into a ball.
The Latin language has the virtue of condensing into one word what might take three in a language such as English. Latin even has a specialised verb, glomero, meaning ‘I roll into a ball’ (from which we get English words like conglomerate and agglomerate). And Glomeris is the scientific name of yet another animal that rolls itself into a ball, and is also called ‘pill’ in vernacular English. It is not a crustacean but a millipede, the ‘pill millipede’, a member of the order Glomerida. As if that wasn’t enough, two different orders of millipede have independently converged on the roll-up pill body. In addition to the order Glomerida, members of the order Sphaerotheriida (Greek ‘spherical beast’) look just like Glomeris and indeed like Armadillidium, except that they are bigger.
Pill woodlouse
Pill millipede
Pill woodlouse (above left) and pill millipede (above right) provide what may be my favourite example – in a strong field – of convergent evolution. They are almost indistinguishable when you see them crawling along, or when they roll into a ball. But the one is a crustacean, related to shrimps and crabs, while the other is a myriapod, related to centipedes. To make sure which is which, I have to turn them over. The crustacean has only one pair of legs per segment, making seven pairs in all. The millipede has many more legs, two pairs per segment. These two deeply different ‘pill’ animals look extremely alike in their surface palimpsest layers because they make their living in the same kind of way and in the same kind of place. Starting from widely separated ancestors they converged, in evolutionary time, on very similar end points.
Giant isopod
The deep palimpsest layers show that one is unmistakeably an isopod crustacean, the other a myriapod. Isopods are an important group of crustaceans, and they include members who grow to alarmingly large size on the sea bottom. We shall refer to them again in the next chapter, which goes to town on crustaceans.
Latin isn’t the only language to impress with its parsimony. The Malay noun pengguling means ‘one who rolls up’ and from it we get the name pangolin. We met the pangolin in the previous chapter. You might mistake it for a large, animated fir cone. It is not closely related to any other mammals but is out in its own order, Pholidota. That name comes from a Greek word meaning ‘covered with scales’, and an alternative English name for pangolin is ‘scaly anteater’. The scales are made of keratin, like hooves and fingernails. They aren’t as hard as the bony armour plates of armadillos.
However, when it comes to glomerising, pangolins perhaps outdo armadillos, pill woodlice, and pill millipedes. According to a report by a biologist on the island of Siberut in Indonesia, a pangolin ran away from him to the top of a steep slope, then formed itself into a ball and rolled down the slope at a speed of about 3 metres per second, twice as fast as a pangolin can run. The witness of this event interpreted the rolling down the hill as a normal response to predation. I reluctantly wonder if it might have been accidental.
There seems to be no doubt as to the effectiveness of rolling up as protection. Lions engage in futile endeavours to penetrate a pangolin’s defence. The pangolin’s enviable insouciance makes one wonder why other hunted animals don’t adopt the same strategy – the tortoise or armadillo strategy – instead of frantically fleeing. I suppose armour is expensive to make, but then so are long, well-muscled, fast-running legs. And it’s not a good argument – though possibly true – that if all antelopes, say, were to jettison speed for armour-plated roll-ups, lions on their side of the evolutionary arms race would come up with a counter-strategy. What might be a better argument is that the first individual antelopes to essay rudimentary, and still inadequate, armour would suffer compared with unencumbered rival antelopes disappearing in a cloud of dust.
Lion thwarted by pangolin
Two of the best-known examples of convergent evolution, too familiar to need detailed illustration yet again, are flight and eyes. The laws of physics allow the possibility of using energy to stay aloft for indefinite periods, and the wing has been independently and convergently invented five times: by insects, pterosaurs, birds, bats, and … human technology.
Eyes have been independently evolved many dozens of times, to nine basic designs. The convergent similarity between the camera, the vertebrate eye, and the cephalopod eye has become almost legendary. Here I’ll just mention that the most revealing difference – the vertebrate retina but not the mollusc one being wired up backwards – is a difference at a deep palimpsest level. This is another way of saying there’s a fundamental difference in their embryology. The vertebrate eye develops mostly as an outgrowth of the brain, while the cephalopod eye develops as an invagination from the outside. That difference lies deep down among the oldest palimpsest layers.
A less familiar example of convergence, compound eyes, have also evolved independently several times. Some bivalve molluscs have a form of compound eye, as do some tube-dwelling annelid worms. These are convergent on each other and on the more highly developed compound eyes of crustaceans, insects, trilobites, and other arthropods. Camera eyes have one lens, which focuses an upside-down image on a retina. The image of a compound eye, if you can call it an image, is the right way up. Think hunting dragonfly, with its pair of large hemispheres, each a cluster of tubes radiating outwards in different directions. Whichever tube sees the target, that’s the direction to fly in order to catch it.
A familiar sight throughout both North and South America is the ‘turkey vulture’. It looks like a vulture, behaves like a vulture, lives the life of a vulture, feeding on carrion that it finds, like a vulture, with a sense of smell keener than is typical among birds. But it is not a vulture. Or rather, it has converged on vulturehood independently of true vultures. But wait, who is to say that Old World vultures are any more ‘true’ than New World turkey vultures? Americans might see the priority differently. Let us call both of them vultures, in enthusiastic recognition of convergent evolution and its impressive power to mislead.
We could settle much the same argument about which are the ‘true’ porcupines. Old World and New World porcupines are both rodents. But within the very large order of rodents, they are not particularly closely related, and they evolved their spiny defences independently. The two pictures show a leopard about to suffer the same punishment from an Old World porcupine as the dog has endured from a New World porcupine.
Contrary to legend, no porcupine shoots its quills. But they do have a quick-release mechanism so that a predator injudicious enough to molest a porcupine comes away with a face full of quills. New World quills prolong the agony by means of backward-facing barbs, which make them difficult to remove. This detail is not shared by the otherwise convergent Old World porcupines but it is convergent, at a much smaller scale, on the barbs of bee stings (American stingers).
Dog after approaching New World porcupine
Leopard approaching Old World porcupine
The sting of a bee, unlike a porcupine quill, is double. There are two barbed blades rubbing against each other with the venom running between them. The two move alternately against each other, sawing their way into the victim. Both are serrated with backward-pointing barbs like those on a New World porcupine quill. The sting is a modified ovipositor, a tube for egg-laying. Porcupine quills are modified hairs. Bees are not the only insects whose ovipositors are serrated. In cicadas (which don’t sting), the serrations, and the bee-like alternate sawing action of the two blades, serve to dig the ovipositor (egg-laying tube) into (for example) a tree, where the eggs are laid.
The sting of a bee, derived from the ovipositor and therefore possessed only by females, is a hypodermic syringe for injecting venom. The hypodermic venom injector has evolved convergently in eleven different animal groups by my count (probably more than once independently in some groups): in insects, scorpions, snakes, lizards, spiders, centipedes, stingrays, stonefish, cone shells, and the hind-leg claw of the male duckbilled platypus. The stinging cells, ‘cnidoblasts’, of jellyfish are miniature harpoons that shoot out on the ends of threads, and inject venom. Among plants, stinging nettles have miniature hypodermic syringes.
The short spikes of hedgehogs are like the long quills of porcupines in being modified hairs. And these too have arisen independently at least three times. There are spiky tenrecs in Madagascar, which look remarkably like hedgehogs although they are not members of the same Order as hedgehogs. They are Afrotheres, related to elephants, aardvarks and dugongs. A third convergence is provided by the spiny anteaters of Australia and New Guinea. Egg-layers, they are as distant from hedgehogs and tenrecs as it is possible to be while still being mammals. They too are covered with spikes, again modified hairs.
We have seen that porcupine quills are a nice example of convergent evolution, independently arisen within the rodents. So-called flying squirrels also arose twice independently in different families of rodents, the true squirrels, and the so-called scaly-tails or anomalures. We know they evolved their gliding habit independently of each other because the closest relatives of both, within the rodents, are not gliders. It’s the same way we know New World and Old World porcupines are convergent, again within the large order of rodents.
Not surprisingly, the gliding skill has evolved convergently in a number of vertebrates. The picture shows four mammal examples, including the two rodents just mentioned. The colugo of the Southeast Asian forests is sometimes called the flying lemur, but it isn’t a lemur (all true lemurs come from Madagascar, though that’s not what makes the colugo a non-lemur) and it doesn’t really fly, although it is perhaps a more accomplished glider than the others in the picture. The sugar glider, although it looks extremely like a flying squirrel, is actually a marsupial from Australasia, one of several ‘flying phalangers’. Despite the startlingly close resemblance between sugar glider and flying squirrel, we know that one is a marsupial and the other a rodent, because of deeper layers of palimpsest. For example, the female phalanger has a pouch, the squirrel a placenta.
1. Colugo
2. Flying squirrel
3. Marsupial sugar glider
4. Anomalure (Not to scale)
The Australian marsupial fauna provides many other examples of convergent evolution, of which perhaps the most famous is the extinct thylacine or Tasmanian wolf, already mentioned. The picture opposite shows a selection of comparisons between Australian marsupials and their placental equivalents in the rest of the world. These include a pair of anteaters and a pair of ‘mice’. The marsupial ‘mole’ of Australia resembles not only the familiar Eurasian mole but also the ‘golden mole’ of South Africa. Also very mole-like, among the rodents there are the zokors of Asia.
All these ‘moles’ independently adopted the same burrowing way of life, all have adapted their hands into powerful spades and all four look pretty alike. So convincing is the convergence that the golden moles were once classified as moles until it was realised that they belong to a radically different branch of (African) mammals, the Afrotheria, together with elephants, aardvarks, and manatees. Eurasian moles, by contrast, are Laurasiatheres, related to hedgehogs, horses, dogs, bats, and whales. Rodent zokors are related to the blind mole rats, who are thoroughly committed to subterranean life and look like moles, but, as you might expect from a rodent, they dig with their teeth rather than their hands. The family tree, overleaf, showing the affinities of four ‘moles’ is quite surprising.
PLACENTAL MAMMALS
MARSUPIALS
Dog
Thylacine
European mole
Marsupial mole
Mouse
Marsupial mouse
Flying squirrel
Sugar glider
Tamandua
Numbat
Independently evolved ‘moles’
Impressive as are the convergences of Australian marsupials with a whole variety of placental mammals, we mustn’t overlook the exceptions. Kangaroos don’t look very like the African antelopes with whom they share a way of life. They easily might have converged. But they didn’t. They diverged, mostly because they early committed themselves to a different gait for travelling fast. I suppose there was a time when the ancestors of either could have adopted the hopping gait of a kangaroo or the galloping gait of an antelope. Both gaits are fast and efficient, a least after many generations of evolutionary perfecting. But once an evolutionary lineage starts down a path like hopping or galloping, it is difficult to change. ‘Commitment’ really is a thing, in evolution. Once a lineage of mammals had advanced some way along the hopping gait path, any mutant that tried to gallop would have been out-competed. Perhaps its front legs were already too short. Conversely, in a lineage that was somewhat committed to galloping, a mutant that tried to hop would clumsily fail. There’s no rule that says placental mammals couldn’t have taken the kangaroo route. Indeed, there are rodents whose ancestors travelled that path very successfully. A colleague teaching zoology at the University of Nairobi said in a lecture that there were no kangaroos in Africa. This was denied by a student who excitedly claimed to have seen a small one. What he had seen was a springhaas or springhare, a rodent that looks and hops just like a wallaby, complete with foreshortened arms and enlarged, counterbalancing tail.
Springhare
If you could witness an ichthyosaur sporting in Mesozoic waves, you’d be irresistibly reminded of dolphins. A classic case of convergent evolution. On the other hand, your time machine might also present to you a plesiosaur. Far from looking like a dolphin or an ichthyosaur, it doesn’t resemble anything else you ever saw. Ichthyosaurs and plesiosaurs are both descended from land reptiles that went back to the sea. But they started out along, and then became ‘committed to’, alternative paths towards efficient swimming ‘gaits’. Ichthyosaurs rediscovered the ancient side-to-side tailbeat of their fish ancestors. They probably passed through a phase resembling the serpentine wavy motion of Galapagos marine iguanas. Plesiosaurs, instead, relied like sea turtles on their limbs, all four of which became huge flippers. Once committed, both ichthyosaurs and plesiosaurs became increasingly dedicated to their respective evolutionary pathways. And ended up looking extremely different.
Convergently evolved animals are not necessarily contemporaries. In North America in the Eocene period there were mole-like subterranean animals, the Epoicotheriids, with mole-like digging hands, not closely related to any living burrowers but belonging to the pangolin family, Pholidota. I’d be surprised if there weren’t dinosaur ‘moles’, but I must confess I don’t know of any. There were smallish dinosaurs such as Oryctodromeus who dug burrows, but I don’t know of any who could be called convergent on moles.
Then there were the so-called ‘false sabretooths’. We’ve already met Smilodon, the sabretooth ‘tiger’, that large, robust and doubtless frightening cat, which went extinct along with most of the American megafauna at the end of the Pleistocene era, only about 10,000 years ago, when man discovered America. What is less well known is that Smilodon was not the only member of the order Carnivora to evolve such terrifying fangs. Thirty million years earlier, spanning the Oligocene epoch, lived a group called Nimravids. The Nimravids were not cats but an older group within the Carnivora, and they independently evolved stabbing canine teeth just like those of Smilodon. Nimravids are sometimes called false sabretooths. False? Tell that to the early horse Mesohippus and the other terrified victims of those giant daggers. Those ‘false’ sabretooths were living, breathing, snarling, pouncing, probably strong-smelling carnivores, to whose victims they would have seemed anything but false. Another extinct group of ‘false sabretooths’, the Barbourofelids, lived in the Miocene epoch, later than the Nimravids but earlier than Smilodon, and convergently occupying the same niche.
‘False’ sabretooth – Nimravid
Given that the Carnivora have endowed us with three independently evolved sabretooths at different times in geological history, we might even feel a little let down if there were no marsupial sabretooth. And sure enough, South America rose to the occasion.
Marsupial sabretooth – Thylacosmilus
The marsupial Thylacosmilus looks to have been nearly as formidable as Smilodon and the other convergent sabretooths of the Carnivora. On the other hand, it was a bit smaller.
Convergences between animals and human technology can be especially impressive, as we saw in the case of the camera and the vertebrate or octopus eye. Though the discovery was originally thought an outrageous hoax, it is now well accepted that bats hunting by night have their own version – ‘echolocation’ – of what submariners have converged upon under the name ‘sonar’ – using echoes of their own sounds to detect targets. Bats are divided into two main groups, the small Microchiroptera, and the large Megachiroptera (‘fruit bats’ and ‘flying foxes’). Microchiropteran bats ‘see’ with their ears. They have highly sophisticated echolocation, good enough to hunt fast-flying insects. The brain pieces together a detailed model of the world, including insect prey, by a highly sophisticated real-time analysis of the echoes of the bats’ own shrieks. When a bat is cruising, its cries just tick over. But when homing in on a moth, which is likely to be taking evasive action, the sounds come out as a rapid-fire stutter like a machine gun. Since each pulse gives the bat an updated picture of the world, machine-gun repetition enables it to cope with a moth’s high-speed twists and turns. The higher the pitch, the shorter the wavelength by definition. And only short wavelengths can resolve a detailed picture. That means ultrasound: too high, mostly way too high, for us to hear. Young people can hear the lower end of the bat’s frequency range. I nostalgically remember them from my youth as sounding like something between a click and a squeak. We can use instruments called bat detectors, which translate ultrasound into audible clicks.
Slightly less well known is the fact that dolphins and other toothed whales (sperm whales, killer whales) do the same thing, also using ultrasound, and they are up there with bats in sophistication. A more rudimentary form of echolocation has also evolved in shrews, and in cave-nesting birds at least twice independently: in South American oilbirds and Asian cave swiftlets (of bird’s-nest soup fame). The birds don’t use ultrasound: their cries are low enough for us to hear. Some megachiropterans also use a less precise form of echolocation, but they generate their clicks with their wings rather than with the voice. This too must be seen as yet another convergent evolution of echolocation. One genus of Megachiroptera echolocates using the voice, like Microchiroptera but not so skillfully. Interestingly, molecular evidence indicates that one group of Microchiroptera, the Rhinolophids, are more closely related to Megachiroptera than they are to other Microchiroptera. This would seem to suggest that the Rhinolophids evolved their advanced sonar convergently with the other Microchiroptera. Either that or the majority of Megachiroptera lost it.
Small bats and toothed whales are in a class of their own. Their sonar is of such high quality that ‘seeing with their ears’ scarcely exaggerates what they do. Echolocation using ultrasound provides them with a detailed picture of their world, which bears comparison with vision. We know this through experimental testing of bats’ ability to fly fast between thin wires without hitting them. I have even published the speculation (probably untestable, alas) that bats ‘hear in color’. I stubbornly maintain that it’s plausible, because the hues that we perceive are internally generated labels in the brain, whose attachment to particular wavelengths of light is arbitrary. When bat ancestors gave up on eyes, substituting echoes for light, the internal labels for hues would have gone begging, left hanging in the brain with nothing to do. What more natural than to commandeer them as labels for echoes of different quality? I suppose you might call it an early exploitation of what some humans know as ‘synaesthesia’.
In one of modern philosophy’s most cited papers, Thomas Nagel didactically asked, ‘What is it like to be a bat?’ One of his points was that we cannot know. My suggestion is that it is perhaps not so very different from what it’s like to be us, or another visual animal like a swallow. Pursuing a point from Chapter 1, both swallows and bats build up an internal virtual reality model of their world. The fact that swallows use light, while bats use echoes, to update the model from moment to moment is less important than the nature and purpose of the internal model itself. This is likely to be similar in the two cases, because it is used for a similar purpose – navigation in real time between obstacles, and detection of fast-moving prey. Swallows and bats need a very similar internal model, a three-dimensional one, inhabited by moving insect targets. Both are champion insect hunters on the wing, swallows by day and then, at nightfall, the bats take over. If my speculation is right, the similarity may extend to the use of colors to label objects in the model, even in the case of bats ‘seeing with their ears’. Incidentally, each swallow eye has two foveas (regions of special acuity – our eyes have only one, which we use for reading etc.), probably one for distance and one for close vision. Instead of bifocal glasses they have bifocal retinas.
The James Webb Telescope presents us with stunning images of distant nebulae, glowing clouds of red, blue and green. Color is used to represent wavelength of radiation. But the colours in the photographs are false. They use color to represent different wavelengths, but they actually lie in the invisible infrared part of the spectrum. And my point is that the brain’s convention for representing visible light of different wavelengths is just as arbitrary. One is tempted to feel dissatisfied by false colour images such as those from the James Webb Telescope: ‘But is that really what it looks like? Is the telescope telling the truth, or are we being fobbed off with false colours?’ The answer is that we are always being ‘fobbed off’ when we look at anything. If you must talk about false colours, everything you ever see – a rose, a sunset, your lover’s face – is rendered in the brain’s own ‘false’ colours. Those vivid or pastel hues are internal concoctions manufactured by the brain as coded labels for light of different wavelength. The truth lies in the actual wavelength of electromagnetic radiation. The perceived hue is a fiction, whether it is the false colour rendering of a James Webb photograph, or whether it is the labels that the brain generates to tag the wavelengths of light hitting the retina. My conjecture about bats ‘hearing in colour’ makes use of the same idea of internally perceived hues being arbitrary labels.
Doctors use ultrasound to ‘look’ through the body wall of a pregnant woman and see a black-and-white moving image of her developing foetus. The computer uses the ultrasound echoes to piece together an image compatible with our eyes. There is anecdotal evidence that dolphins pay special attention to pregnant women swimming with them. It seems plausible that they are doing with their ears what doctors do with their instruments. If this is so, they could presumably also ‘see’ inside female dolphins and detect which ones are pregnant. Might this skill be useful to male dolphins choosing mates? No point inseminating a female who is already pregnant.
Bats and dolphins evolved their echo-analysing skills independently of each other. In the family tree of mammals, both are enveloped by relatives who don’t do echolocation. A strong convergence, and another powerful demonstration of the power of natural selection. And now for a point that’s especially telling for the genetic book of the dead. There’s a type of protein called prestin, which is intimately involved in mammal hearing. It’s expressed in the cochlea, the snail-shaped hearing organ in the inner ear. As with all biological proteins, the exact sequence of amino acids in prestins is specified by DNA. And, also as is usual, the DNA sequence is not identical in different species. Now here’s the interesting point. If you construct a family tree of resemblance based on the genome as a whole, whales and bats are far apart, as you’d expect: their ancestors have been evolving independently of one another since way back in the age of dinosaurs. If, however, you ignore all genes except the prestin gene – if you construct a tree of resemblance based on prestin sequences alone – something remarkable emerges. Dolphins and small bats cluster together with each other. But small bats don’t cluster together with non-echolo-cating large bats, to whom they are much more closely related. And dolphins don’t cluster together with baleen whales, which, although related to them, don’t echolocate. This suggests that SOF could read the prestin gene of an unknown animal and infer whether it (more precisely its ancestors) lived and hunted in conditions where ultrasonic sonar would be useful: night, dark caves, or other places where eyes are useless, such as the murky water of the Irrawaddy river or the Amazon. I’d like to know whether the two echolocating bird species have bat-like prestins.
This finding on bats and dolphins – the specific resemblance of their prestin genes – strikes me as a pattern for a whole field of future research on the genetic book of the dead. Another example concerns flight surfaces in mammals. Bats fly properly, and marsupial flying phalangers glide, using stretched flaps of skin that catch the air. There’s a specific complex of genes, shared by both bats and marsupial phalangers, which is involved in making the skin flaps. It will be interesting to know whether the same genes are shared by the other gliding mammals that we met earlier in this chapter, so-called flying lemurs and the two groups of rodents that independently evolved the gliding habit.
It would be nice to look in the same kind of way at those animals who have returned from land to water – of which whales are only the most extreme example, along with dugongs and manatees. Do returnees to water have genes in common that are not shared by non-aquatic mammals? What other features do they share? Many aquatic mammals and birds have webbed feet. If our hypothetical SOF is presented with an unknown animal who has webbed feet, she can safely ‘read’ the feet as saying, ‘Water in the recent ancestral environment.’ But that’s obvious. Can we be systematic in our search for less obvious signals of water in the genetic book of the dead? How many other features are diagnostic of aquatic life? Are there some shared genes, such as we saw in the case of prestin for sonar, and skin flaps in bats and sugar gliders? There are probably lots of shared features buried deep in an aquatic animal’s physiology and genome. We have just to find them. We can get a sort of negative clue by looking at genes that were made inactive when terrestrial animals took to the water. Just as humans have a large number of smell genes inactivated (see here), whale genomes contain several inactivated genes, whose inactivation has been interpreted as beneficial when diving to great depths.
We could proceed along the following lines. We borrow from medical science the technique known as GWAS (genome-wide association study). The idea of GWAS is lucidly and conversationally explained by Francis Collins, former Director of the Human Genome Project, as follows:
What you do for a genome-wide association study is find a lot of people who have the disease, a lot of people who don’t, and who are otherwise well matched. And then, searching across the entire genome … you try to find a place where there is a consistent difference. And if you’re successful – and [you’ve] got to be really careful about the statistics here, so that you don’t jump on a lot of false positives – it allows you to zero in on a place in the genome that must be involved in disease risk without having to guess ahead of time what kind of gene you’re going to find.
Substitute ‘lives in water’ for ‘disease’, and ‘species’ for ‘people’, and you have the procedure I am here advocating. Let’s call it ‘Interspecific GWAS’ or IGWAS.
Gather a large number of mammals known to be aquatic. Match each one with a related mammal (the more closely related the better) who lives on land, preferably in dry conditions. We might start with the following list of matched pairs, and the list could be extended.
Water vole
Vole
Water shrew
Shrew
Desman
Mole
Platypus
Echidna
Water tenrec
Land tenrec
Otter
Badger
Seal
Wolf
Yapok
Opossum
Polar bear
Brown bear
To do the IGWAS, you would now look at the genomes of all the animals and try to pinpoint genes shared by the left-hand column and not by the right-hand column. Until all those animals have had their genomes sequenced, and until mathematical techniques are up to the task, proceed with a non-genomic version of IGWAS as follows. Go to work taking measurements of all the animals. Measure all the bones. Weigh the heart, the brain, the kidneys, the lungs, etc., all these weights being expressed relative to total body weight (to correct for absolute size, which is unlikely to be of much interest). By the same token, the bone measurements should be expressed as a proportion of something, just as, in the chelonian example of Chapter 3, the bone lengths were expressed as a proportion of total arm length. Measure the body temperature, blood pressure, the concentrations of particular chemicals in the blood, measure everything you can think of. Some of the measurements might not be continuously varying quantities like centimetres or grams: they might be ‘yes or no’, ‘present or absent’, ‘true or false’.
Feed all the measurements into a computer. And now for the interesting part. We want to maximise the discrimination between aquatic mammals and their terrestrial opposite numbers. We want to discover which measurements discriminate them, pull them apart. At the same time, we want to identify those features that unite all aquatic mammals, however distantly related from each other. Webbing between the toes will presumably emerge as a good discriminator, but we want to find the non-obvious discriminators, biochemical discriminators, ultimately gene discriminators. Where genomic comparisons are concerned, the GWAS methods already developed for medical purposes will serve. A possible graphic method is a version of the triangular plot of tortoise and turtle limbs that we saw in Chapter 3. Another graphic method is drawing pedigrees with genetic convergences coloured in.
A refinement of IGWAS might order species along an ecological dimension. You could, perhaps, string mammals out along a dimension of aquaticness, from whales and dugongs at one extreme to camels, desert foxes, oryxes, and gundis at the other. Seals, otters, yapoks and water voles would be intermediate. Or we might explore a dimension of arboreality. We might conclude that a squirrel is a rat who has moved a measurable distance along the dimension of arboreality. Are moles, golden moles and marsupial moles situated at one extreme on a dimension of fossoriality. Could we distribute birds along a dimension from flightless cormorants and emus who never fly, at one extreme, to albatrosses at the other, or, even more extreme, to swifts, who even copulate on the wing? Having identified such ‘dimensions’, could we look for trends in gene frequency as you move along from one extreme to the other. I can immediately foresee alarming complications. The dimensions would interact with other dimensions, and we’d have to call in experts with mathematical wings to fly through multi-dimensional spaces. My own sadly amateur ventures, limited to three dimensions, and using computer simulation rather than mathematics, are in my book Climbing Mount Improbable, especially the chapter called ‘The Museum of All Shells’.
A group at Carnegie Mellon University in Pittsburgh performed a model example of what I call (they don’t) IGWAS. What they studied was not aquaticness but hairlessness in mammals. Most mammals are hairy, and all had hairy ancestors, but if you survey the mammal family tree you notice that hairlessness pops up sporadically among unrelated mammals. See the diagram, which shows a few of the sixty-two species whose genomes were examined.
Sporadic distribution of hair loss among mammals
Whales, manatees, pigs, walruses, naked mole rats, and humans have all lost their hair more or less completely (yellow names in the diagram). And, which is important, independently of each other in many cases. We can tell this by looking at the hairy closer relatives from among whom they sprang. You remember that echolocating bats and echolocating whales had something else in common – their prestin gene. Do the genomes of the naked species have a gene for hairlessness that they share with each other? The answer is literally no. But only literally. The truth is equally interesting. It turns out that we and other naked species still retain the ancestral genes that make hairs. But the genes have been disabled. And disabled in different ways. What is convergent is the fact of being disabled, but the details are not shared. Incidentally, we again have here a problem for creationists. If an intelligent designer wished to make a naked animal, why would he equip it with genes for making hair and then disable them? Chapter 3 mentions the similar example of the human sense of smell: the olfactory sense genes of our mammal ancestors still lurk within us, but they have been turned off.
One of my favourite examples of convergent evolution is that of weakly electric fish. Two separate groups of fish, Gymnotids in South America and Gymnarchids in Africa, have independently and convergently discovered how to generate electric fields. They have sense organs all along the sides of the body, which can detect distortions that objects in the environment cause in the electric fields. It is a sense of which we can have no awareness. Both groups of fish use it in murky water where vision is impossible. There’s just one difficulty. The normal undulating movements typical of fish fatally compromise the analysis of the electric fields measured along the body. It is necessary for the fish’s body to maintain a rigid stance. But if their body is rigid, how do they swim? By means of a single longitudinal fin traversing the whole length of the body. The body itself, with its row of electrical sensors, stays rigid, while the single longitudinal fin alone performs the sinuous movements typical of fish locomotion. But there’s one revealing difference. In the South American fish, the longitudinal fin runs along the ventral surface, while in the African fish it runs along the back. In both groups of fish, the undulating waves can be thrown into reverse: the fish swim backwards and forwards with apparently equal facility.
The ‘duck bill’ of the platypus and the huge, flat ‘paddle’ sticking out of the front end of the paddlefish (Polyodontidae) are both covered with electrical sensors, convergently and independently evolved. In this case the electric fields they pick up are generated, inadvertently, by the muscles of their prey. There is a long-extinct trilobite that also had a huge paddle-like appendage like that of the paddlefish. Its paddle was studded with what look like sense organs, and it seems probable that this represents yet another convergence.
A ringed plover’s eggs and chicks lie out on the ground, defenceless except for their camouflage. A fox approaches. The parent is much too small to put up any kind of resistance. So it does an astonishing thing. It attempts to lure the predator away from the nest by offering itself as a bigger prize than the nest. It limps away from the nest, pretending to have a broken wing, simulating easy prey. It flutters pathetically on the ground, wings outstretched, sometimes with one wing stuck incongruously in the air. There’s no assumption that it knows what it is doing or why it is doing it (although it may). The minimal assumption we need make is that natural selection has favoured ancestors whose brains were genetically wired up to perform the distraction display, and perfect it over generations. Now, why tell the story in this chapter on convergent evolution? It’s because the broken wing display has arisen not once but many times independently in different families of birds. The diagram on the following page is a pedigree of birds, wrapped around in a circle so it fits on the page. Birds who perform the broken wing display are coloured in red, those who don’t in blue. You can see that the habit is distributed sporadically around the pedigree, a lovely example of convergent evolution.
My final example of convergence will lead us into the next chapter. More than 200 species, belonging to thirty-six different fish families, practise the ‘cleaner’ trade. They remove surface parasites and damaged scales from the bodies of larger ‘client’ fish. Each individual cleaner fish has its own cleaning station, and its own loyal clients who return repeatedly to the same ‘barber’s shop’ on the reef. This site tenacity is important in keeping the benefit exclusively mutual: the cleaner eats the parasites and worn-out scales from the skin of particular client fish, and the client refrains from eating its particular benefactor. Without individual site fidelity, and therefore repeat visits, clients would have no incentive to refrain from eating the cleaner – after being cleaned, of course. Sparing a cleaner would benefit fish in general, including competitors of the sparer. Natural selection doesn’t ‘care’ about general benefit. Quite the contrary. Natural selection cares only about benefit to the individual and its close relations, at the expense of competitors. A bond of individual loyalty between particular cleaner and particular client therefore really matters, and it is achieved by site tenacity. Some cleaners even venture inside the mouth of a client to pick its teeth – and survive to repeat the service on the client’s next visit. Cleaner fish advertise their trade and secure their safety by a characteristic dance, often enhanced by a striped pattern – the fishy equivalent of the striped pole insignia of a human barber’s shop. This constitutes a safe-conduct pass.
Broken wing display
The remarkable ‘broken wing display’ crops up again and again in different bird groups (shown in red). Striking testimony to the power of natural selection.
The interesting point for this chapter is that the cleaner habit has evolved many times convergently, not only many times independently in fish but many times in shrimps too. As before, the client fish abide by the covenant and refrain from eating their cleaner shrimps, in just the same respectful way as for cleaner fish. In many cases, cleaner shrimps sport a similar stripe, the ‘barber’s pole’ insignia. It is to the benefit of all that all the ‘barber’s pole’ badges should look similar.
When swimming in the sea, you would be well advised to steer clear of the sharp-toothed jaws of the moray eel. Yet here is a shrimp, calmly picking its teeth. Note, yet again, the red stripe or ‘barber’s shop pole’, telling the moray, ‘Don’t eat me, I’m your special cleaner. You and I have a mutual relationship. You’ll need me again.’ Does the shrimp feel fear as it trustingly enters those formidable jaws? Does some equivalent of ‘trust’ pulsate through its cephalic ganglion? I doubt it, but not everyone would agree. Do you?
Moray eel and cleaner shrimp
Not only has the habit evolved independently – convergently – in fish and shrimps. It has evolved convergently many times within shrimps, just as it has many times within fish. Even within one family of shrimps, the Palaemonidae, the cleaner trade is practised by sixteen different species, having evolved within the Palaemonidae five times independently. Here’s how we know the five evolutions were independent of each other. The method again serves as a model for how we ever know instances of evolution are independent of each other. Look at the family tree of the Palaemonidae, constructed with the aid of molecular genetic sequencing. It contains sixty-eight species of shrimp. Those species that practise the fish-cleaning trade have a little fish symbol by them. There are sixteen species of palaemonid cleaner shrimps. But many of the sixteen cannot be said to have evolved the habit independently. For example, the three species of Urocardella are all cleaners, but the picture warns us against counting them as independent: they probably inherited it from their common ancestor.
Six members of the genus Ancyclomenes are cleaners, but again we must make the conservative assumption that they inherited it from their common ancestor – and that the habit has been lost in A.aqabai, A.kuboi, A.luteomaculatus, and A.venustus. Using this conservative approach, we conclude that the cleaning habit evolved independently in five palaemonid genera but not in all species of those five genera. And the story doesn’t end with the Palaemonidae. Two other families of shrimps not shown in the diagram, the Hippolytidae (see moray eel picture above) and the Stenopodidae, also have many species of cleaner.
The Cambridge palaeontologist Simon Conway Morris has treated convergent evolution more vividly and thoroughly than anyone else. In his wittily written Life’s Solution he points out that convergent evolution is commonly sold as amazing, astounding, uncanny, etc., but there is no need for this. Far from being especially amazing, it’s exactly what we should expect of natural selection. Convergent evolution is, nevertheless, great for confounding armchair philosophers and others who underestimate the power of natural selection and the magnificence of its productions. In addition to 110 densely packed pages of massively researched endnotes and references to the biological literature, Life’s Solution has three indexes: a general index, a name index and – this must surely be unique – a ‘convergences index’. It runs to five double-column pages and around 2,000 examples of convergence. Of course, not all of them are as impressive as the pillbugs, the moles, the gliders, the sabretooths, or the fish-cleaners but even so …
Independent evolution of cleaners
Convergent evolution can be so impressive, it makes you wonder how we know the resemblance really is convergent. That’s the power of natural selection, the immense yet subtle power that underpins the whole idea of the genetic book of the dead. Pill woodlouse and pill millipede, alike as two pills, how do we know one is a crustacean, the other a distant myriapod? There are numerous tell-tale clues. The deep layers of the palimpsest are never completely over-written. The glyphs of history keep breaking through. And, if all else fails, molecular genetics cannot be denied.
Convergence of animals with widely separated histories is one manifestation of the power of selection to write layer upon layer of the palimpsest. Another is its converse: evolutionary divergence from a common historic origin, natural selection seizing a basic design and moulding and twisting it into an often bizarre range of functionally important shapes. The next chapter goes there.
6 Variations on a Theme
As we saw in Chapter 3, molecular comparison conclusively shows that whales are located deep within the even-toed ungulates, the artiodactyls. By ‘located deep within’, I mean something very specific and surprising. It’s worth repeating. We’re talking about much more than just a shared ancestor, with the whales going one way, and the artiodactyls the other. That would not have been surprising. ‘Deep within’ means that some artiodactyls (hippos) share a more recent ancestor with whales than they share with the rest of the artiodactyls whom they much more strongly resemble. This has been known for more than twenty years, but I still find it almost incredible, so overwhelming is the submersion under surface layers of palimpsest. Of course, this doesn’t mean whales’ ancestors were hippos or even resembled hippos. But whales are hippos’ closest living relatives.
What is it that’s so special about whales, so special that new writings in their book of the dead so comprehensively obliterated almost every trace of that earlier world, of grazing prairies and galloping feet, which must lie buried far down in the palimpsest? How did the whales manage to diverge so completely from the rest of the artiodactyls? How were they able so comprehensively to escape their artiodactyl heritage?
The answer probably lies in that word ‘escape’. Cattle, pigs, antelopes, sheep, deer, giraffes, and camels are relentlessly disciplined by gravity. Even hippos spend significant amounts of time on land, and indeed can accelerate their ungainly bulk to an alarming speed. The land-dwelling artiodactyl ancestors of whales had to submit to gravity. In order to move, land mammals must have legs stout enough to bear their weight. A land animal as big as a blue whale would need legs half way to Stonehenge pillars, and it’d have a hard time surviving, with heart and lungs smothered suffocatingly by the body’s own weight. But in the sea, whales shook off gravity’s tyranny. The density of a mammal body is approximately that of water. Gravity never goes away, but buoyancy tames it. When their artiodactyl ancestors took to the water, whales shed the need for leggy support, and the fossil evidence beautifully lays out the intermediate stages.
A major milestone marks the point where, like dugongs and manatees but unlike seals and turtles, whales gave up returning to land even to reproduce. That was the final release from gravity, as buoyancy totally took over. Whales were free to grow to prodigious size, literally insupportable size. A whale is what happens when you take an ungulate, cut it adrift from the land and liberate it from gravity. All manner of other modifications followed in the wake of the great emancipation, and they richly defaced the ancient palimpsest. Forelegs became flippers, hind limbs disappeared inside and shrank to tiny relics, the nostrils moved to the top of the head, two massive horizontal flukes – lobes stiffened not by bone but by dense fibrous tissue – sprouted sideways to form the propulsive organ. Numerous profound alterations of physiology and biochemistry allowed deep diving, and hugely prolonged intervals between breaths. Whales switched from a (presumed) herbivorous diet to one dominated by fish, squid, and – in the case of the baleen whales – filtered shoals of krill in lavish quantities.
Fish, too, are allowed by buoyancy to adopt bizarre shapes (see pictures here), which gravity on land would forbid. In the case of teleost (bony as opposed to cartilaginous) fish, the buoyancy is perfect, owing to that exquisite device, the swim-bladder, buried deep within the body. By manipulating the amount of gas in the swim-bladder, the fish is able to adjust its specific gravity and achieve perfect equilibrium at whatever happens to be its preferred depth at any time.
I think that’s what makes a home aquarium such a restful furnishing for a room. You can dream of drifting effortlessly through life, as a fish drifts through water in perpetual equilibrium. And it is the same hydrostatic equilibrium that frees fish to assume such an extravaganza of shapes. The leafy sea dragon trails clouds of glorious fronds, and you feel you could almost identify the species of wrack that those fronds mimic. You must peer deep between them to discern that they are parts of a fish: a modified sea horse – which is itself a distorted caricature of the ‘standard fish’ design of more familiar cousins such as trout and mackerel.
Most predatory fish actively seek and pursue prey, and this expends a considerable proportion of the energy obtained from the food caught. Angler fish, of which there are several hundred species sitting on the sea bottom, save energy by luring prey to come to them. The anglers themselves are superbly camouflaged. A fishing rod (modified fin spine) sprouts from the head. At its tip is a lure or bait, which the angler fish waves around in a tempting manner. Unsuspecting prey are attracted to the bait, whereupon the angler opens its enormous mouth and engulfs the prey. Different species of angler favour different baits. With some it resembles a worm, and it jiggles about plausibly as the angler waves its rod. Angler fish of the dark deep sea harbour luminescent bacteria in the tip of the rod. The resultant glowing lure is very attractive to other deep-sea fish, and invertebrate prey such as shrimps. Convergently, snapping turtles rest with their mouth open, wiggling their tongue like a worm, as bait for unsuspecting prey fish.
Sea horses and angler fish are extreme exponents of the adaptive radiation of teleost fish. They also, in their different ways, sport unusual sex lives. The sex life of angler fish is nothing short of bizarre. Everything I said in the previous paragraph applies to female angler fish only. The males are tiny ‘dwarf males’, hundreds of times smaller than females. A female releases a chemical, which attracts a dwarf male. He sinks his jaws into her body, then digests his own front end, which becomes buried in the female’s body. He becomes no more than a small protuberance on her, housing male gonads from which she extracts sperm when she needs to. It is as though she becomes a hermaphrodite, except that ‘her’ testes possess a different genotype from her own, having invaded from outside in the form of the dwarf male locked into her skin.
Lionfish
Weedy sea dragon
Marlin
Leafy sea dragon
Trumpet fish
Sunfish
Gulper eel
Seahorse
Puffer
Sloane’s viper fish
Ghost pipefish
Angler fish
Freed by buoyancy from the constraints of gravity, fish were able to evolve an astonishing variety of shapes
Many species of fish are livebearers – females get pregnant like mammals and give birth to live young. Sea horses are unusual in that it’s the male who gets pregnant, carries the young in a belly pouch, and eventually gives birth to them. Do you wonder, then, how we define him as male? Throughout the animal and plant kingdoms, the male sex is easily defined as the one that produces lots of small gametes, sperms, as opposed to fewer, larger, eggs.
Adaptive radiation means evolutionary divergence fanning out from a single origin. It is seen in an especially dramatic way when new territory suddenly becomes available. When, 66 million years ago, a celestial catastrophe cleared 76 per cent of all species from the planet, the stage was wide open for mammalian understudies to step into the dinosaurs’ vacated costumes. The subsequent adaptive radiation of mammals was spectacular. From the small, burrowing creatures who survived the devastation, probably by hibernating in safe little underground bunkers, a comprehensive range of descendants, ranging hugely in size and habit, appeared in surprisingly quick time.
On a smaller scale and a much shorter timescale, a volcanic island can spring up suddenly (suddenly by the standards of geological time) through volcanic upwelling from the bottom of the sea. For animals and plants it is virgin territory, barren, untenanted, open to exploitation afresh. Slowly (by the standards of a human lifetime) the volcanic rock crumbles and starts to make soil. Seeds fly in on the wind, or are transported by birds and fertilised with their droppings. From being a black lava desert, the island greens. Winged insects waft in, and tiny spiders parachuting under floating threads of silk. Migrating birds are blown off course, land for recuperation, stay, reproduce; their descendants evolve. Fragments of mangrove drift in from the mainland, and the occasional tree uprooted by a hurricane. Such freak raftings carry stowaways – iguanas, for instance. Step by accidental step, the island is colonised. And then descendants of the colonists evolve, rapidly by geological standards, diversifying to fill the various empty niches. Diversification is especially rich in archipelagos, where driftings between islands happen more frequently than from the mainland to the archipelago. Galapagos and Hawaii are textbook examples.
A volcano is not the only way new virgin territory for evolution can open up. A new lake can do it too. Lake Victoria, largest lake in the tropics and larger than all but one of the American Great Lakes, is extremely young. Estimates range from 100,000 years to a carbon-dated figure of only 12,400 years. The discrepancy is easily explained. Geological evidence shows that the lake basin formed about 100,000 years ago, but the lake itself has dried up completely and refilled several times. The figure of 12,400 years represents the age of the latest refilling, and therefore the age of the current lake in its large geography. And now, here is the astonishing fact.
There are about 400 species of Cichlid (pronounced ‘sicklid’) fish in Lake Victoria, and they are all descended from probably as few as two founder lineages that arrived from rivers within the short time that the lake has existed. The same thing happened earlier in the other great lakes of Africa, the much deeper Lakes Tanganyika and Malawi. Each of the three lakes has its own unique radiation of Cichlid fishes, different from, but parallel to, the others.
Nimbochromis livingstonii
Lamprologus lemairii
Here’s a slightly macabre example of this parallelism. In Lake Malawi (where I spent my earliest bucket-and-spade beach holidays), there is a predatory fish called Nimbochromis livingstonii. It lies on the bottom of the lake pretending to be dead. It even has light and dark blotches all over its body, giving the appearance of decomposition. Deceived into boldness, small fish approach to nibble at the corpse, whereupon the ‘corpse’ suddenly springs into action and devours the small fish. This hunting technique was thought to be unique in the animal kingdom. But then exactly the same trick was discovered in Lake Tanganyika, the other great Rift Valley lake. Another Cichlid fish, Lamprologus lemairii, has independently, convergently, hit upon the same death-shamming trick. And it has the same blotchy appearance, suggestive of death and decay. In both lakes, adaptive radiation independently hit upon the same somewhat gruesome way of getting food. Along with dozens of other ways of life, independently discovered in parallel in the two similar lakes.
My old friend, the late George Barlow, vividly described the three great lakes of Africa as Cichlid factories. His book, The Cichlid Fishes, makes fascinating reading. The Cichlids have so much to teach us about evolution in general and adaptive radiation in particular. Each of the three great lakes has its own, independently evolved radiation of several hundred Cichlid species. All three lakes tell the same story of explosive Cichlid evolution, yet the three histories unfolded entirely independently. All three began with a founder population of very few species. Each of the three followed a parallel evolutionary course of massive radiation into a huge variety of ‘trades’ or ways of life – the same great range of trades being independently discovered in all three lakes.
You might think the oldest lake would have the most species. After all, it’s had the longest time to evolve them. But no. Lake Tanganyika, easily the oldest at about 6 million years, has only (only!) 300 species. Victoria, a baby of only 100,000 years, has about 400 species. Lake Malawi, intermediate in age at between 1 and 2 million years, has the largest species count, probably around 500, although some estimates exceed 1,000. Moreover, the size of the radiation seems unrelated to the number of founder species. The huge radiations in Victoria and Malawi trace back substantially to only one lineage of Cichlids, the Haplochromines. The relatively venerable Lake Tanganyika’s approximately 300 species appear to stem from twelve different founder lineages, of which the Haplochromines are only one.
What all this suggests is that young Lake Victoria’s dramatic explosion of species is the model for all three lakes. All three probably took only tens of thousands of years to generate several hundred species. After the explosive beginning, the typical pattern is probably to stabilise the number, or it may even decrease, such that the final number of species is not correlated with the age of the lake, or with the number of founder species. The Cichlids of Lake Victoria show how fast evolution can proceed when it dons its running shoes. We cannot expect that such an explosive rate is typical of animals in general. Think of it as an upper bound.
And when you work it out, even Lake Victoria’s feat is not quite so surprising as first appears. Although the lake in its present form is only some 12,400 years old, I’ve already mentioned that a lake filled the same shallow basin 100,000 years ago. In the intervening years it has largely dried up several times and refilled, the latest such episode occuring with the refill of 12,400 years ago. Lake Malawi shows how dramatically these lake levels can fall and rise. Between the fourteenth and nineteenth centuries, the water level was more than 100 metres lower than today. Unlike Lake Victoria, however, it came nowhere close to drying up altogether. In its Rift Valley chasm, it is nearly ten times as deep as Victoria. In shallow Lake Victoria, as each drying cycle occurred, the lowering of the water level would have left numerous ponds and small lakes, these becoming reunited at the next iteration of the refill cycle. The temporary isolation of the fish trapped in the residual ponds and small lakes enabled them to evolve separately – no gene flow between ponds. At the next refill of the cycle, they were reunited, but by then they would have drifted apart genetically, too far to interbreed with those who had been stranded in other ponds. If this is correct, the drying/refilling alternation provided ideal conditions for speciation (the technical term for the evolutionary origin of a new species, by splitting of an existing species). And it means that, from an evolutionary point of view, we could regard the true age of Lake Victoria as 100,000 years, not 12,400. Still very young.
Given 100,000 years to play with, what sort of interval between speciation events would yield 400 species, starting, hypothetically, with a single founding species? Is 100,000 years long enough? Here’s how a mathematician might reason: a back-of-the-envelope calculation, making conservative assumptions throughout, to be on the safe side. There are two extremes, two bounds bracketing the possible rate of speciation, depending on the pattern of splitting. The most prolific pattern (an improbable extreme) is where every species splits into two, yielding two daughter species which, in turn, split into two. This pattern yields exponential growth of species numbers. It would take only between eight and nine speciation cycles to yield 400 species (29 is 512). An interval of 11,000 years between speciations would do the trick. The least prolific pattern (also an improbable extreme) is where the founder species ‘stays put’ and successively throws off one daughter species after another. This would require far more speciation events, about 400, to reach the tally of 400 species: a speciation event every 250 years. How to estimate a realistic intermediate between these two extremes? A simple average (arithmetic mean) gives an estimate of between 5,000 and 6,000 years between speciations, which is enough time. Our mathematician, however, might be more cautious and recommend the geometric mean (multiply the two numbers together and take the square root). One reason to prefer it is that it captures the stronger influence of an occasional very bad year. This more conservative estimate asks for an interval of about 1,600 years between speciations. Somewhere between the two estimates is plausible, but let’s bend over backwards to be cautious and use the estimate of 1,600 years. Cichlid fish typically reach sexual maturity in under two years, so let’s again be conservative and assume a two-year generation time. Then we’d need about 800 fish generations between speciation events, in order to generate 400 species in 100,000 years. Eight hundred generations is enough for plenty of evolutionary change.
How do I know 800 generations is plenty of time? Again, mathematicians can do back-of-the-envelope calculations to assist intuition. One calculation that I like was done by the American botanist Ledyard Stebbins. Imagine that natural selection is driving mouse-sized animals towards larger size. Stebbins, too, bent over backwards to be conservative, by assuming a very weak selection pressure, so weak that it could not be detected by scientists working in the field, trapping mice and measuring them. In other words, natural selection in favour of larger size is assumed to exist but to be so slight and subtle that it is below the threshold of detectability by field researchers. If the same undetectably weak selection pressure were maintained consistently, how long would it take for the mice to evolve to the size of an elephant? The answer Stebbins calculated was about 20,000 generations, the blink of an eye by geological standards. Admittedly, it’s a lot more than our 800 generations, but we weren’t talking about anything so grandiose as mice turning into elephants. We were only talking about Cichlid fishes changing enough to be incapable of interbreeding with other species. Moreover, Stebbins’s assumptions, like ours, were conservative. He assumed a selection pressure so weak that you couldn’t measure it. Selection pressures have actually been measured in the wild, for example on butterflies. Not only are they easily detectable, they are orders of magnitude stronger than the sub-threshold, under-the-radar pressure assumed by Stebbins. I conclude that 100,000 years is a comfortably long time in Cichlid evolution, easily enough time for an ancestral species to diversify into 400 separate species. That’s fortunate, because it happened!
Incidentally, Stebbins’s calculation is an instructive antidote to sceptics who think geological time is not long enough to accommodate the amount of evolutionary change we observe. His 20,000 generations to wreak the change from mouse to elephant is so short that it would ordinarily not be measurable by the dating methods of geologists. In other words, a selection pressure too weak to be detectable by field geneticists is capable of yielding major evolutionary change so fast that it could look instantaneous to geologists.
The crustaceans are another great group of mostly aquatic animals with spectacular evolutionary radiations, from much more ancient common sources. In this case, it is the modification of a shared anatomy that impresses. Rigid skeletons permit movement only if built up of hinged units, bones in the case of vertebrates, armoured tubes and casings in the case of crustaceans and other arthropods. Because these bones and tubes are rigid and articulated, there is a finite number of them, each one a unit that can be named and recognised across species. The fact that all mammals have almost the same repertoire of nameable bones (206 in humans) makes it easy to recognise evolved differences as distortions of each named bone: ulna, femur, clavicle, etc. The same is true of crustacean skeletal elements, with the bonus that, unlike bones, they are externally visible.
The great Scottish zoologist D’Arcy Thompson took six species of crab and looked at just one unit of the skeleton, the main portion of the body armour, the carapace, of each.
Geryon
Corystes
Chorinus
Scyramathia
Lupa
Paralomis
He arbitrarily chose one of the six, it happened to be Geryon (far left), and drew it on a rectangular grid. He then showed that he could approximate the shape of each of the other five, simply by distorting the grid in a mathematically lawful way. Think of it as drawing one crab on a sheet of stretched rubber, then distorting the rubber sheet in mathematically specified directions to simulate five other shapes. These distortions are not evolutionary changes. The six species are all contemporary. No one species is ancestral to any other, they share ancestors who are no longer with us. But they show how easily changes in embryonic development (altered gradients of growth rates, for instance) can yield an illuminating variety of crustacean form with respect to one part of the exoskeleton. D’Arcy Thompson did the same thing with many other skeletal elements including human and other ape skulls.
Of course, bodies are not drawn on anything equivalent to stretched rubber. Each individual develops afresh from a fertilised egg. But changes in growth rates, of each part of the developing embryo, can end up looking like the distortions of stretched rubber. Julian Huxley applied D’Arcy Thompson’s method to the relative growth of different body parts in the developing embryo. Such embryological changes are under genetic control, and evolutionary changes in gene frequencies generate evolutionary variety, again looking like stretched rubber. And of course it isn’t just the carapace. The same kind of evolutionary distortion is seen in all the elements of the crustacean body (and the bodies of all animals but often less obviously). You can see how the same parts are present in each specimen, just emphasised to different degrees. The differential emphasis is achieved by different growth rates in different parts of the embryo.
Crustaceans are exceedingly numerous. With characteristic wit, the Australian ecologist Robert May said, ‘To a first approximation, all species are insects,’ yet it has been calculated that there are more individual copepods (crustacean water fleas) than there are individual insects in the world. The painting opposite, by the zoologist Ernst Haeckel (1834–1919), Darwin’s leading champion in Germany, is a dazzling display of the anatomical versatility of the copepods.
Wondrous copepods from Ernst Haeckel’s Art Forms in Nature
Mantis shrimp
Here’s a typical adult crustacean, a mantis shrimp. Well, mantis shrimps (Stomatopods) are typical with respect to their body plan, which, together with their colourful beauty, is why I’ve chosen one for this purpose. But they include some formidable customers who are far from typical in one alarming respect. They pack a punch, literally. With vicious blows from club-like claws, they smash mollusc shells in nature, while in captivity the blow from a large smasher, travelling as fast as a small-calibre rifle bullet, will shatter the glass of your aquarium tank. The energy released is so great that the water boils locally and there is a flash of light. You don’t want to mess with a mantis shrimp, but they’re a wonderful example of the diverse modification of the basic crustacean body plan.
Mantis shrimps are not to be confused with the (literally) stunning ‘pistol shrimps’ or ‘snapping shrimps’ (Alpheidae), who in their way also beautifully illustrate the diversity of crustacea. These have one enlarged claw, somewhat bigger than the other. They snap the enlarged claw with terrific force, generating a shock wave – a violent pulse of extreme high pressure immediately followed by extreme low pressure in its wake. The shock wave stuns or kills prey. The noise is among the loudest heard in the sea, comparable to the bellows and squeaks of large whales. Muscles are too slow to generate high-speed movement such as the snapping claws of pistol shrimps or the punching clubs of mantis shrimps (or indeed the jump of a flea). They store energy in an elastic material or spring, and then suddenly release it – the catapult or bow-and-arrow principle.
Crustacea dazzle with diversity. But it is a constrained diversity. To repeat the point, which is the reason I chose crustaceans for this chapter, you can in every species easily recognise the same parts. They are connected to each other in the same order, while differing hugely in shape and size. The first thing you notice about the basic crustacean body plan is that it is segmented. The segments are arrayed from front to rear like a goods train with trucks (American freight train with wagons or cars). The segmentation of centipedes and millipedes is even more obviously train-like because most of their segments are the same. A mantis shrimp or a lobster is like a train whose trucks are the same in a few respects (wheels, bogies, and coupling hooks, say) but different in other ways (cattle wagons, milk tanks, timber carriers, etc.).
Crustaceans in their evolution achieve astonishing variety by changing the trucks over evolutionary time, while never losing sight of the train. Varied as they are, the segments of a mantis shrimp are still visibly a train built to the same pattern as any other crustacean, each bearing a pair of limbs that fork at the tip. The claw of a crab or lobster is a conspicuous example of the fork. As you move from front to rear of the animal, the paired appendages consist of antennae, various kinds of mouth parts, claws, then four pairs of legs. Move backwards further, and the segments of a lobster or mantis shrimp’s abdomen each have small, jointed appendages called swim-merets underneath, on both sides, each often ending in a little paddle. In a lobster or, even more so, a crab, the segments of the thorax and head are hidden beneath a shared cover, the carapace. But their segmentation is betrayed by the appendages, walking legs in the case of four of them, antennae, large claws and mouth parts at the front end. The rear end of the abdomen, the guard’s van (American caboose) of the train, has a special pair of flattened appendages called uropods. When I first visited Australia, I was intrigued to see, laid out in a buffet, what they call bay bugs. These have what look like uro-pods at the front end as well as the rear, a sort of crustacean version of Doctor Dolittle’s Pushmi-Pullyu, but with two rear ends instead of two heads. This is not all that surprising, as we shall now see.
The segmentation of arthropods and vertebrates was once thought to have evolved independently. No longer, and thereby hangs a fascinating tale, a tale that is true too of other segmented animals such as annelid worms. Just as the segments are arrayed in series from front to rear like a train, so the genes controlling the segments are arrayed in series along the length of a chromosome. This revolutionary discovery overturned the whole attitude to zoology that I had learned as a student, and I find it wonderful. To pursue the railway analogy, there’s a train of gene trucks in the chromosome to parallel the train of segment trucks in the body.
It’s been known for more than a century that mutant fruit flies can have a leg growing where an antenna ought to be. That mutation is called antennapedia for obvious reasons, and it breeds true. There are other dramatic mutations in fruit flies, for example bithorax, which has four wings like normal insects, instead of the two-winged pattern that gives flies their name, Diptera. These major mutations are all explained by changes in the sequentially arranged genes in the ‘chromosome train’. When I first saw that bay bug in a Great Barrier Reef restaurant, I immediately wondered whether bay bugs had originally evolved by a mutation similar to antennapedia, in this case duplicating uropods at the front end of the animal.
This kind of effect has been neatly shown by Nipam Patel and his colleagues. They work on a marine crustacean called Parhyale, belonging to the Amphipod order. I remember being fascinated by the hundreds of small amphipods in the cold stream on our farm, in the course of which my parents dug out a pool for us to swim. The swarms of exuberantly jumping ‘sandhoppers’ that we so often encounter on beaches are another familiar example. We met iso-pods, in the flattened shape of ‘pill bugs’, in the previous chapter. Amphipods are different. They are flattened left to right rather than back to belly. And, in Parahyale and many others, their appendages are far from all the same. Some of their legs point in what seems to be the ‘wrong’ direction. Three of the ‘trucks’ appear to be ‘coupled’ up backwards (red shading in left picture on the next page). Patel and his colleagues, by means of ingenious manipulations of the genes controlling the trucks of the train, were able to change the three reversed segments, coupling the trucks so that all the limbs faced in the same direction (right picture). The way this works is that the three backwards segments are replaced by duplicates of the three segments in front of them. The Patel group achieved equally interesting manipulations of other segments but the work, though fascinatingly ingenious, would take us too far afield.
ILLUSTRATION: KALLIOPI MONOYIOS
We vertebrates too are segmented, but in a different way. This is obvious in fish, and it remains pretty clear in our ribs and vertebral column. Snakes carry it to an extreme – sort of like centipedes but with internal ribs instead of external legs. We now understand the embryological mechanism whereby segments are multiplied up. Surprisingly, actually rather wonderfully, it has turned out to be pretty much the same in vertebrates and arthropods. Hence, we understand how it is that different snake species evolve radically different numbers of vertebrae ranging from around 100 to more than 400 – compared to our thirty-three. Vertebrae, whether or not they sprout ribs, all have similar coupling mechanisms to the neighbouring ‘trucks of the train’, and all have similar blood vessels, and sensory and motor nerves, connected to the spinal cord, which passes through them. As I just mentioned, one of the most revolutionary discoveries of recent zoology is that the embryological mechanisms underlying segmentation in arthropods and vertebrates, deep in the lower levels of their palimpsests, are tantalisingly similar. Once again, the truly beautiful fact is that in both groups, genes are laid out along chromosomes in the same order as the segments that they influence.
Although crustaceans all follow the segmented plan boldly written in the depths of the palimpsest, the ‘trucks’ vary so extravagantly that the simile of the train can become rather strained. Sometimes many of the segments join together to form a singular body, as in crabs. Often the appendages sprouting from the segments vary spectacularly, ranging from the formidable claws near the front of a lobster, or the punching clubs of a mantis shrimp, to the swimmerets arrayed under the abdomen. Crustaceans range in size from ‘water fleas’ at less than 1 millimetre to the Japanese spider crab Macrocheira with a limb span that can reach 3 metres (10 feet). Frightening as this creature might be to meet, it is harmless to humans. Imagine the handshake of a lobster, or the punch of a mantis shrimp, that size!
Japanese spider crab
Crabs can be thought of as lobsters with a truncated tail (abdomen) curled up under the main body, so you don’t see it unless you upend the animal. The crab abdomen bears a passing resemblance to the ape/human coccyx, both being made of a handful of segments from an ancestral tail squashed up. Hermit crabs are strictly not crabs, but belong in their own group (Anomura) within the crustacea. Their abdomen is not squashed up underneath them as in true crabs, but soft and curled round to one side, to fit the discarded mollusc shells that hermit crabs inhabit. The process by which they choose their shells, and compete with one another for favoured shells, is fascinating in its own right. But that’s another story. In this chapter they serve as yet another illustration of the wonderful diversity of crustaceans.
The larvae of crustaceans show the group’s diversity at least as gloriously as the adults. But still the basic train design is palpable throughout. Perhaps even more dramatically than in the case of adult crustaceans, it is as though natural selection pulled, pushed, kneaded, or distorted the various segments of the body with wild abandon. Different species of crustacean pass through nameable larval stages, free-living animals in their own right, often leading a very different life from the adults – as caterpillars live very differently from butterflies among the insects. The zoea is one such larval type. It is the last stage before the adult, in crabs, lobsters, crayfish, shrimps, bay bugs, and their kind – the decapod crustaceans.
Overleaf is a page full of assorted zoeas to show how easily the basic crustacean plan can be stretched and bent around in evolution, as though made of modelling clay. What I take away from these exquisite little creatures is that all have the same parts, they just vary the relative sizes and shapes of those parts. They all look like distorted versions of each other. That’s what evolutionary diversification is all about, and the crustacea show it as plainly as any animal group. You can match up the corresponding parts in all the species, and can clearly see how the different species have pulled, stretched, twisted, swelled, or shrunk the same parts in different ways over evolutionary time. It is wondrous to behold, you surely agree.
Crustacean larvae. Always the same parts, yet pulled and pushed in different directions
Zoeas may look a little like the adults they are to become. But they need to survive in a very different world, usually the world of plankton, and their bodies are versatile enough to evolve into all sorts of unlikely distortions – written in surface layers of the palimpsest. Many of them sport long spikes, presumably to make them difficult to swallow. The impressive spikes of the planktonic zoea at top middle are nowhere to be seen in the typical adult crab it is to become. Truth be told, the adult in this case is not easily seen at all under the sea urchin that it habitually carries around on its back – presumably to gain protection via the urchin’s own spikes. Notice the long, prominent abdomen of the larva, with its easily discerned segments. As with all crabs, the adult abdomen is neither long nor prominent but tucked discreetly under the thorax.
An earlier larval stage than the zoea, found in most crustacean life cycles, is the nauplius larva. Unlike zoeas, which bear some sort of resemblance to the adult they will become, naupliuses have an appearance all their own. There’s another larval stage possessed by some crustaceans, the cyprid larva, presumably so called because it resembles the adult of a water flea called Cypris. Perhaps the adult Cypris is an example of the overgrown larva phenomenon, which is a fairly common way for evolution to progress. Below is the cyprid larva of a member of the rather obscure crustacean sub-class, Facetotecta.
Facetotectan larva
This larva is unmistakeably crustacean, with a head shield, and abdominal segments bearing typically crustacean forked appendages. From 1899, when the larvae were first discovered, until 2008, nobody knew what adult facetotectans looked like. And they still have never been seen in the wild. What happened in 2008 was that a group of experimentalists succeeded in persuading larvae to turn into a precursor of the adult. They did it by means of hormone treatment. The subtitle of their paper is ‘Towards a solution to a 100-year-old riddle’. The adults turn out to be soft, unarmoured, slug-like or worm-like creatures with no visible segments and no appendages, presumably parasites, although nobody knows who their victims are. You wouldn’t know, to look at them, that they are crustaceans at all. This experiment recalls a similar one by Julian Huxley with axolotls in 1920. Axolotls are vertebrates, members of the Amphibia. They look like tadpoles; indeed they are tadpoles, but sexually mature tadpoles, and they reproduce. They evolved from larvae who would once have turned into salamanders. The adult stage of their life history was cut off during their evolution, as the larvae became sexually capable. By treating them with thyroid hormone, Julian Huxley succeeded in turning them into the salamanders that their ancestors once were. This experiment may have inspired his younger brother Aldous Huxley to write his novel After Many a Summer, in which an eighteenth-century aristocrat discovered how to cheat death – and developed, 200 years later, into a shaggy, long-armed ape humming a Mozart aria. We humans are ‘larval’ apes!
Those slug-like facetotectans are yet another manifestation of crustacean diversity. They must be descended from adults who had segments and limbs like any respectable crustacean. But the most characteristically crustacean scripts of the palimpsest have been almost completely obliterated by parasitic over-writing, while being retained in the larva. Degenerative evolution of this kind is common in parasites hailing from many parts of the animal kingdom. Within the crustacea, it is also shown to an extreme in certain members of the barnacle family, though not the typical barnacles that encrust rocks at the seaside and prick your bare feet when you walk on them.
As a boy on a seaside holiday, I remember being frankly incredulous when my father told me barnacles are really crustaceans. I thought they were molluscs because, well, they look like molluscs. Nothing like crustaceans, anyway, until you look carefully inside. The barnacles that cling close to rocks look like miniature limpets, while goose barnacles look like mussels on stalks. So how do we know they are really crustaceans? Look inside. Or see Darwin’s own drawing above and you find a shrimp-like creature lying on its back and sweeping the water with its comb-like limbs to filter out swimming morsels of food. As we have by now come to expect, the larvae of barnacles are more unmistakeably crustacean than the adults. Before the adult settles down to its sedentary permanence, it is a free-swimming larva in the plankton. On the left is the nauplius larva of Semibalanus, a small rock barnacle with, for comparison, the nauplius larva of a shrimp, Sicyonia.
Barnacle larva
Shrimp larva
Barnacles don’t encrust only rocks. To a barnacle, a whale would seem like a gigantic mobile rock. Not surprisingly, some barnacles make their home on the surface of whales, and there are species of barnacle who live nowhere else. Others ride on crabs, and some of them, especially Sacculina, evolved into the most extreme examples of divergence from normal crustacean form. They moved, in evolutionary time, from the outside of the crabs to the inside, and became internal parasites bearing no apparent resemblance to a barnacle – or even any kind of animal. Parasites often evolve in a direction that could fairly be called degeneration, and Sacculina is an extreme example of this. I shall return to it in the final chapter.
There are many groups of animals that I could have chosen to illustrate evolutionary divergence and variation on a theme. Fish and crustaceans do it perhaps more spectacularly than any other groups, and I chose especially the larvae of crustaceans, partly because, living in the plankton as most of them do, they are less familiar than adult lobsters, crabs, and prawns. I regret that in this book I have been able to show only a small number of them. See the splendid Atlas of Crustacean Larvae, published by Johns Hopkins University Press, for the full and amazing range of diversity that these mesmerising little creatures display. Sir Thomas Browne (1605–82) was unaware of them when he wrote the following, about bees, ants, and spiders, but crustacean larvae might have moved him to even greater eloquence.
Ruder heads stand amazed at those prodigious pieces of nature, Whales, Elephants, Dromedaries and Camels; these I confess, are the Colossus and Majestick pieces of her hand but in these narrow Engines there is more curious Mathematicks, and the civilitie of these little Citizens more neatly sets forth the wisdome of their Maker.
7 In Living Memory
The most recent scripts, those in the top layer of the palimpsest, are those written during the animal’s own lifetime. I said that the genes inherited from the past can be seen as predicting the world into which an animal is going to be born. But genes can predict only in a general way. Conditions change on a timescale faster than the generational turnover with which natural selection can cope. Many details are usefully filled in during the animal’s own lifetime, mostly by memories stored in the brain, as opposed to the genetic book of the dead, in which ‘memories’ are written in DNA. Like gene pools, brains store information about the animal’s world, information that can be used to predict the future, and hence aid survival in that world. But brains can do it on a swifter timescale. Strictly speaking, where learning – indeed, this whole chapter – is concerned, we are talking not about the genetic book of the dead but about the non-genetic book of the living. However, as we shall see, naturally selected genes from the past prime the brain to learn certain things rather than others.
The gene pool of a species is sculpted by the chisels of natural selection, with the result that an individual, programmed as it is by a sample of genes drawn from the well-carved gene pool, tends to be good at surviving in environments that did the carving: that is, an averaged set of ancestral environments. An important part of the body’s equipment for survival is the brain. The brain – its lobes and crevices, its white matter and grey matter, its bewildering byways of nerve cells and highways of nerve trunks – is itself sculpted by natural selection of ancestral genes. The brain is subsequently changed further by learning, during the animal’s lifetime, in such a way as to improve yet further the animal’s survival. ‘Sculpting’ might not seem so appropriate a word here. But the analogy between learning and natural selection has impressed many, not least BF Skinner, a leading – if controversial – authority on the learning process.
Skinner specialised in the kind of learning called operant conditioning, using a training apparatus that later became known as the Skinner Box. It’s a cage with an electrically operated food dispenser. An animal, often a rat or a pigeon, gets used to the idea that food sometimes appears in the automatic dispenser. Built into the wall of the box is a pressable lever or a peckable key. Pressing the lever or key causes food to be delivered, not every time but on some automatically scheduled fraction of occasions. Animals learn to operate the device to their advantage. Skinner and his associates have developed an elaborate science of so-called operant conditioning or reinforcement learning. Skinner Boxes have been adapted to a wide variety of animals. I once saw a film of a rotund gourmand, in a specially reinforced Skinner Box, noisily exercising the lever-bashing skill of his bulbous pink snout. I found it endearing, and I hope the pig enjoyed it as much as I enjoyed the spectacle.
You can train an animal to do almost anything you like, by operant conditioning, and you don’t have to use the automated Skinner Box apparatus. Suppose you want to train your dog to ‘shake hands’, that is, politely raise his right front paw as if to be shaken. Skinner called the following technique ‘shaping’. You watch the animal, waiting until he spontaneously makes a move that you perceive as being slightly in the right direction: an incipient, tentative, upward movement of the right front paw, say. You then reward him with food. Or perhaps not with food but with a signal such as the sound of a ‘clicker’, which he has previously been taught to associate with a food reward. The clicker is known as a secondary reward or secondary reinforcement, where the food is the primary reward (primary reinforcement). You then wait until he moves his right front paw a little further in the right direction. Progressively, you ‘shape’ his behaviour closer and closer to the target you have chosen, in this case ‘shaking hands’. You can use the same shaping technique to teach a dog to do all manner of cute tricks, even useful ones like shutting the door when there’s a cold draught and you are too lazy to get out of your armchair. It is elaborations of the same shaping technique that erstwhile circus trainers employed to teach bears and lions to do undignified tricks.
I think you can see the analogy between behaviour ‘shaping’ and Darwinian selection, the parallel that so appealed to Skinner and many others. Behaviour-shaping by reward and punishment is the equivalent of shaping the bodies of pedigree dogs by artificial selection – domestic breeding. The gene pools of pedigree cattle, sheep, and cats, of racehorses and greyhounds, pigs and pigeons, have been carefully sculpted by human breeders over many generations to improve running speed, milk or wool yield, or in the case of dogs, cats, and pigeons, aesthetic appeal according to various more-or-less bizarre standards. Darwin himself was an enthusiast of the pigeon fancy, and he devoted an early chapter of On the Origin of Species to the power of artificial selection to modify domestic animals and plants.
Now, back to shaping in Skinner’s sense. The animal trainer has a particular end result in mind, such as handshaking in a dog. She waits for spontaneous ‘mutations’ (please note well the quotation marks) of behaviour thrown by an individual animal and selects which ones to reward. As a consequence of the reward, the chosen spontaneous variant is then ‘reproduced’ by the animal itself in the form of a repetition. Next, the trainer waits for a new ‘mutant’ (again please don’t ignore the quotation marks) extension of the desired behaviour. When the dog spontaneously goes a little further in the desired direction of the handshake, she rewards him again. And so on. By a careful regimen of selective rewards, the trainer shapes the dog’s behaviour progressively towards a desired end.
The analogy with genetic selection is evident and was expounded by Skinner himself. But so far, the analogy is with artificial selection. How about natural selection? What role does reinforcement learning play in the wild, where there are no human trainers? Does the analogy with reward learning extend from artificial selection to natural selection. How does reward learning improve the animal’s survival?
Darwin bridged the gap from domestic breeding to natural selection with his great insight that human breeders aren’t necessary. Human selective breeders – let’s call them gene pool sculptors – are replaced by natural sculptors: the survival of the fittest, differential survival in wild environments, differential success in attracting mates and vanquishing sexual rivals, differential parenting skills, differential success in passing on genes. And just as Darwin showed that we don’t need a human breeder, the analogy with learning does without a human trainer. With no human trainers, animals in the wild learn what’s good for them and shape their behaviour so as to improve their chances of survival.
‘Mutation’ consists of spontaneous trial actions that might be subject to ‘selection’ – i.e. reward or punishment. The rewards and punishments are doled out by nature’s own trainers. When a hen scratches the ground with her feet, the action has a good chance of uncovering food of some kind, perhaps a grub or a seed. And so ground-scratching is rewarded, and repeated. When a squirrel bites the kernel of a nut, it’s hard to crack unless held at a particular angle in the teeth. When the squirrel spontaneously discovers the right angle of attack, the nut cracks open, the squirrel is rewarded, the correct alignment of the teeth on the nut is remembered and repeated, and the next nut is cracked more quickly.
Much depends on the rewards that nature doles out. Food is not the only reward that we can use, even in the lab. Once, for a research project that I needn’t go into, I wanted to train baby chickens to peck differently coloured keys in a Skinner Box. There were reasons not to use food as reward, so I used heat instead. The reward was a two-second blast from a heat lamp, which the chicks found agreeable, and they readily learned to peck keys for the heat reward. But now we need to face the question, what, in general, do we mean by ‘reward’? As Darwinians, we must expect that natural selection of genes is ultimately responsible for determining what an animal treats as rewarding. It’s not obvious what will be rewarding, however obvious it might seem to us because we are animals ourselves.
We may define a reward as follows. If a random act by an animal is reliably followed by a particular sensation and if, in consequence, the animal tends to repeat the random act, then we recognise that sensation (presence of food or warmth or whatever it is) as a reward by definition. If a Skinner Box delivered not food or heat but an attractive and receptive member of the opposite sex, I have no doubt that it would – at least under some circumstances – fit the definition of a reward: an animal in the right hormonal condition would learn to press a key to obtain such a reward. A mother animal, cruelly deprived of her child, would learn to press a key to restore access. And the child would learn to press a key to obtain access to its lost mother. I know of no direct evidence for any of those guesses, nor for my conjecture that a beaver would treat access to branches, stones, and mud suitable for dam-building as a reward by the above definition. And a crow in the nesting season would define access to twigs as a reward. But as a Darwinian, in all those cases I make the prediction with a modicum of confidence.
Brain scientists are able to implant electrodes painlessly in the brains of animals, through which they can stimulate the brain electrically. Normally they do this in order to investigate which parts of the brain control which behaviour patterns. The experimenter controls an animal’s behaviour by passing weak electric currents. Stimulate a chicken’s brain here, and the bird shows aggressive behaviour. Stimulate a rat’s brain there, and the rat lifts its right front paw. The neurologists James Olds and Peter Milner conceived a variant of the technique. They handed the switch over to the rat. By pressing a lever, rats were able to stimulate their own brain. Olds and Milner discovered particular areas of the brain where self-stimulation by rats was highly rewarding: the rats appeared to become addicted to lever-pressing. Not only did electrical stimulation in these brain regions fulfil the definition of a reward. It did so in a big way. When the electrodes were inserted in these so-called pleasure centres, rats would obsessively press the switch, to the extent of unfortunately neglecting other vital activities. They would sometimes press the lever at a rate of 7,000 presses per hour, would ignore food and receptive members of the opposite sex and go for the lever instead, would run across a grid delivering electric shocks in order to get at the lever. They would press the lever continually for twenty-four hours until the experimenters removed them for fear they’d die of starvation. The experiments have been repeated on humans with similar results. The difference is that humans could verbalise what it felt like:
A sudden feeling of great, great calm … like when it’s been winter, and you have just had enough of the cold, and you go outside and discover the first little shoots and know that spring is finally coming.
Another woman (and you have to wonder whether the experiment was approved by an ethics committee)
quickly discovered that there was something erotic about the stimulation, and it turned out that it was really good when she turned it up almost to full power and continued to push on her little button again and again … she often ignored personal needs and hygiene in favor of whole days spent on electrical self-stimulation.
Rat addict
It seems plausible that natural selection has wired up animal brains in such a way that external stimuli or situations that are good for the animal (which will vary from species to species) are internally connected to the ‘pleasure centres’ discovered by Olds and Milner.
Punishment is the opposite of reward. If an action is reliably followed by a stimulus X and, as a consequence, the animal becomes less likely to repeat the action, then X is defined as a punishment. In the laboratory, psychologists sometimes use electric shock as punishment. More humanely (I guess) they use a ‘time out’ – an interval during which the animal is denied access to reward. Dog trainers (the practice is frowned upon by many experts, rightly in my opinion) sometimes smack an animal as punishment. When I was at boarding school (and this practice is now not only frowned upon but illegal) my friends and I were from time to time caned by the headmaster, hard enough (astonishing as it now seems) to leave bruises that took weeks to heal (and were admired at bath-time like battle scars). What my offences were I have now forgotten, but I’m sure I didn’t forget while I was still at the school and within range of Slim Jim and Big Ben, the two canes in the headmaster’s quiver. My probability of repeating the offence undoubtedly decreased. Therefore, beatings were punishments by definition, as well as by the intention of the headmaster.
In nature, bodily injury is perceived as painful. If an action is followed by pain, the probability of repeating that action goes down. Not only is that how we define punishment: it also explains what pain is for, in the Darwinian sense. Injury often presages death and hence failure to reproduce. Therefore, the nervous system defines bodily injury as painful.
Sometimes pain is endured when offset by reward. We’ve already seen that rats will endure painful electric shock to get to the self-stimulation lever. The punishment of a bee sting may be offset by the reward of honey. The taste of honey is such an intense reward that many animals, including bears, honey badgers, raccoons, and human hunter-gatherers, are prepared to endure the pain for the sake of it. Rewards and punishments trade off against each other, just as mutually opposing natural selection pressures trade off against each other.
The Darwinian interpretation of pain as a warning not to repeat the preceding action has ethical implications. In our treatment of non-human animals, on farms and hunting fields, in slaughterhouses and bullrings, we are apt to assume that their capacity to suffer is less than ours. Are they not less intelligent than we are? Surely this means they feel pain, if at all, less acutely than us? But why should we assume that? Pain is not the kind of thing you need intelligence to experience.
The capacity to feel pain has been built into nervous systems as a warning, an aid to learning not to repeat actions that caused bodily damage and might next time lead to death. So, if a species is less intelligent, might its pain need to be more agonising, rather than less? Shouldn’t humans, being cleverer, get away with less painful pain in order to learn not to repeat the self-harming action? A clever animal, you might think, could get away with a mild warning, ‘Er, probably a good idea not to do that again, don’t you think?’ Whereas a less intelligent animal would need the sort of dire warning that only excruciating pain can deliver. How should this affect our attitude towards slaughterhouses and agricultural husbandry? Should we not, at very least, give our animal victims the benefit of the doubt? It’s a thought, to put it at its mildest!
Rewards and punishments, pleasure and pain, are so familiar and obvious to us as human animals that you probably wonder why I am labouring the topic in this chapter. Here is where things start to become less obvious and more interesting. The brain’s choice of what shall constitute reward and what punishment is not fixed in stone. It is ultimately determined by genetic natural selection. Animals come into the world equipped with genetically granted definitions of reward and punishment. These definitions have been made by natural selection of ancestral genes. Any sensation associated with an increased probability of death will become defined as painful. A dislocated limb in the wild dramatically increases the probability of death. And it is intensely painful, as I recently and very vocally testified, all the way to the hospital. It has certainly made me take great care to avoid risking a repeat. Copulation increases the probability of reproduction, and genetic selection has consequently made the accompanying sensations pleasurable – which means rewarding. It has been suggested, with support from rat experiments and from the self-stimulating woman mentioned above, that sexual pleasure is directly linked to the ‘pleasure centres’ discovered by Olds and his colleagues. Presumably other sensations, too, could be so linked by natural selection.
I conjecture that by artificial selection you could breed a race of pigeons who enjoy listening to Mozart but dislike Stravinsky. And vice versa. After many generations of selective breeding, perhaps spread over several human lifetimes, the birds would be genetically equipped with a definition of reward such that they would learn to peck a key that caused a recording of Mozart to be played, and would learn to peck a key that caused a recording of Stravinsky to be switched off. And of course, the experiment would be incomplete unless we also bred a line of pigeons who treated Mozart as punishment and Stravinsky as reward. Let’s not get pedantic as to whether it is really Mozart that they’d treat as rewarding. The learned preference would probably generalise from Mozart to Haydn! The only point I am trying to make is that the definitions of what is rewarding and what is punishing are not carved in stone. They are carved in the gene pool and therefore potentially changeable by selection.
As a corollary, I conjecture that, by artificial selection, you could (though I wouldn’t wish to, and it might take an unconscionable number of generations) breed a race of animals who regarded what had previously been pain as rewarding. By definition, it would no longer be pain! It would be cruel to release them into their species’ natural environment because, of course, they would be unfitted to survive there – that’s the whole point. But the mere fact that they enjoy what normal members of their species would call pain is not cruel – because, however hard it is for us to imagine, at least within the confines of my thought experiment, they enjoy it! Anyway, the more interesting conclusion is that, in a state of nature, it is natural selection that determines what is reward and what is punishment. My thought experiment was devised to dramatise that conclusion.
Experimental psychologists have long known that you can train an animal to treat as a reward something that previously had neutral value for the animal. As mentioned above, it’s called secondary reinforcement, and an example is the clicker used by dog trainers. But secondary reinforcement is not what I’m talking about here, and I really want to emphasise that. I’m not talking about secondary reinforcement, but about genetically changing the very definition of what constitutes primary reinforcement. I conjecture that we could achieve it by breeding, as opposed to training. I called it a conjecture because the experiment has not, as far as I know, been done. I’m now talking about selectively breeding animals in such a way as to change their own genetically instilled definition of what constitutes a primary reward in training. To repeat my suggestion above, I predict that by artificial selection you could in principle breed a race of animals who would treat bodily injury as rewarding.
Douglas Adams carried the point to a wonderful comedic reductio in The Restaurant at the End of the Universe. Zaphod Beeblebrox’s table was approached by a large bovine creature, who announced himself as the dish of the day. He explained that the ethical problem of eating animals had been solved by breeding a species that wanted to be eaten and was capable of saying so. ‘Something off the shoulder, perhaps, braised in a white wine sauce?’
Birds don’t naturally listen to human music, so my Mozart/Stravinsky flight of fancy may seem implausible. But do they have a music of their own? A respected ornithologist and philosopher named Charles Hartshorne suggested that we should regard birdsong as music, appreciated aesthetically by the birds themselves. He may not have been wrong, as I shall soon argue.
The role of learning and genes in the development of birdsong has been intensively studied, especially by WH Thorpe, Peter Marler, and their colleagues and students. Many birds learn to imitate the song of their father or other members of their own species. Spectacular feats of mimicry by the likes of mynahs and lyre birds are an extreme. In addition to mimicking other species such as kookaburras (‘laughing jackass’), lyre birds have been recorded by David Attenborough giving remarkably convincing imitations of car alarms, camera shutters (with or without a motor drive), the chainsaws of lumberjacks and the mixed noises of a building site. I have even heard it said, but have failed to verify it, that lyre birds can distinctly mimic Nikon versus Canon camera shutters. Such virtuoso mimics incorporate an amazing variety of such sounds in an ample repertoire.
This raises the question of why many songbirds have large repertoires in the first place. Individual male nightingales can sport more than 150 recognisably distinct songs. Admittedly that’s an extreme, but the general phenomenon of song repertoires demands an explanation. Given that song serves to deter rivals and attract mates, why not stick to one song? Why switch between alternatives? Several hypotheses have been proposed. I’ll mention just my favourite, the ‘Beau Geste’ hypothesis of John Krebs.
In the adventure yarn of that name by PC Wren, an outnumbered unit of the French Foreign Legion was beleaguered in a desert fort, and the commander beat off the opposing force with a spectacular bluff.
As each man fell, throughout that long and awful day, [the commander] had propped him up, wounded or dead, set the rifle in its place, fired it, and bluffed the Arabs that every wall and every embrasure and loophole of every wall was fully manned.
Krebs’s hypothesis is that the bird with a large repertoire is pretending his territory is already occupied to the full. He is, as it were, mimicking the sounds that would emerge from an area if it were already overpopulated with too many members of his species. This deters rivals from attempting to set up their territory in the area. The more densely populated an area is, the less will it benefit an individual to settle there. Above a certain critical density, it pays an individual to leave and seek territory elsewhere, even an otherwise inferior territory. So, by pretending to be many nightingales, an individual nightingale seeks to persuade others to find a different place to set up his territory. In the case of lyre birds, the sound of a chainsaw is just another addition to the repertoire, the size of which conveys the message: ‘Go away, there’s no future for you here, the place is fully occupied.’
Virtuoso impressionists like lyre birds, mynahs, parrots, and starlings are outliers. Probably they are just manifesting, in extreme form, the normal way young birds learn their species song – imitating their fathers or other species members. The point of learning the correct species song is to attract mates and intimidate rivals. And now we return to our discussion of the definition of a reward: how natural selection defines what will be treated as reward and what punishment.
In an experiment by JA Mulligan, three American song sparrows (Melospiza melodia) were reared by canaries in a soundproof room so that they never heard the song of a song sparrow. When they grew up, all three produced songs that were indistinguishable from those of typical wild song sparrows. This shows that song sparrow song is coded in the genes. But it is also learned. In the following special sense. Young song sparrows teach themselves to sing, with reference to a built-in template, a genetically installed idea of what their song ought to sound like.
What’s the evidence for this? It is possible surgically, under anaesthetic and I trust painlessly, to deafen birds. This has been done, with both song sparrows and the related white-crowned sparrows, Zonotrichia leucophrys. If birds of either species are deafened as adults, they continue to sing almost normally: they don’t need to hear themselves sing. As adults, that is. If, however, they are deafened when three months old, too young to sing, their song when they reach adulthood is a mess, bearing little resemblance to the correct song. On the template hypothesis, this is because they have to teach themselves to sing, matching their random efforts against the template of correct song for the species. There’s an interesting difference between the two species. Whereas the song sparrow never needs to hear another bird sing – its template is innate – the white-crowned sparrow makes a ‘recording’ of white-crowned sparrow song, early in life, long before it starts to develop its own song. Once the template is in place, whether innate as in the song sparrow or recorded as in the white-crowned, the nestlings then use it to teach themselves to sing.
Doves and chickens push this to an extreme: they don’t need to listen to themselves, ever. Ring dove (also known as barbary dove) squabs, who have been surgically rendered completely deaf, later develop vocalisations that are just like those of intact doves. That the behaviour is innate is further testified by the fact that hybrid doves coo in a way that is intermediate between the parental species’ coos. As we shall see in Chapter 9, young crickets (nymphs), before they achieve their final moult to become adults, can artificially be induced to display nerve-firing patterns identical to their species song patterns, even though nymphs never sing. And hybrid crickets have a song that is intermediate between the two parental species.
But I want to get back to the sparrows. As we have seen, they teach themselves to sing by listening to their own random babblings, and repeating those fragments that are rewarded by a match to a template – whether the template is genetically built-in (song sparrow), or a ‘recording’ (white-crowned sparrow) remembered from infancy. Did you notice this means that a sound that matches the template is a reward by our definition? We have identified a new kind of reward to add to food and warmth. The song template is a much more specialised kind of reward. It’s easy to see how food (relief of hunger pangs) and warmth (relief of cold discomfort) would be general, non-specific rewards. Indeed, psychologists of the early twentieth century delighted in reducing all rewards to one simple formula, which they called ‘drive reduction’. Hunger and thirst were seen as examples of ‘drives’, analogous to forces driving the animal. A particular pattern of sounds, complicated and characteristic enough to be recognised, by ornithologists and birds alike, as belonging to one species and one species alone, is a reward of a very different kind from generalised drive-reduction. And, I would personally add, of a much more interesting kind. As a student I tried to read up that rat psychology literature, and I’m sorry to admit that I found it rather boring compared to the zoology literature on wild animals.
The ethologist Keith Nelson once gave a conference talk with the title ‘Is bird song music? Well, then, is it language? Well, then, what the hell is it?’ It isn’t language: not rich enough in information, and it doesn’t seem to be grammatical in the sense of possessing a hierarchical nesting of ‘clauses’ enclosing ‘sub-clauses’. Hartshorne, as I mentioned previously, thought it was music, and I think there’s a sense in which he was right. I believe we can make a case that birds have an aesthetic sense, which responds to song. I think there’s also a sense in which it works like a drug. In what follows, I am drawing on a pair of papers that I wrote jointly with John Krebs some years ago, about animal signals generally. We were critically responding to a then prevalent idea that animal signals function to convey useful information from the sender to the recipient, for the mutual benefit of both. For example, ‘I am a male of the species Luscinia megarhynchos, I am in breeding condition, and I have a territory over here.’ The gene’s-eye view of evolution, then quite novel, did not sit well with ‘mutual benefit’. Krebs and I followed the gene’s-eye view to a more cynical view of animal signals, substituting the idea of manipulation of the receiver by the signaller. ‘You are a female of the species Luscinia megarhynchos. COME HITHER! COME HITHER! COME HITHER!’
When an animal seeks to manipulate an inanimate object, it has only one recourse – physical power … But when the object it seeks to manipulate is itself another live animal there is an alternative way. It can exploit the senses and muscles of the animal it is trying to control … A male cricket does not physically roll a female along the ground and into his burrow. He sits and sings, and the female comes to him under her own power.
Now, you might object, surely the female should respond to male song in this way only if it benefits her. But we regarded the relationship between signaller and signallee as an arms race, run in evolutionary time. Perhaps she does put up some sales-resistance. But that provokes the male, on the other side of the arms race, to up the ante: increase the intensity of his signal. And now we come to another strand to the argument, which Krebs and I advanced in the second of our two papers. This concerns what we called ‘mind-reading’. Any animal in a social encounter can benefit itself by predicting (behaving as if predicting) the behaviour of another. There are all kinds of give-away clues. If a male dog raises his hackles, this is an involuntary indicator of an aggressive mood. Responding appropriately to such give-aways is what we dubbed ‘mind-reading’. Humans can become quite adept at mind-reading in this sense, making use of such cues as shifty eyes or fidgety fingers. And now, to bring the argument full circle, an animal who is the victim of a mind-reader can exploit the fact of being mind-read, in such a way as to render inappropriate the very word ‘victim’. A male, for instance, might manipulate a female by ‘feeding’ her mind-reading machinery, perhaps with deceptive cues. This is just to say that where victimhood is concerned, manipulation is not a one-way street. Mind-reading turns the tables. And then manipulation potentially turns them back again, against the mind-reader.
On this view animal signals, to repeat, evolve as an arms race between mind-reading and manipulation, an arms race between salesmanship and sales-resistance. In those cases where the sender benefits from being mind-read and the receiver benefits from being manipulated, we suggested that the ensuing signal should shrink to a ‘conspiratorial whisper’. Why escalate a signal when there is no push-back. Conversely – the opposite of a conspiratorial whisper – loud, conspicuous, vivid signals will arise where the recipient does not ‘want’ to be manipulated. In such cases the arms race, in evolutionary time, escalates towards exaggeration on the part of the sender, to combat increased ‘sales-resistance’ on the part of the receiver.
Why, you might wonder, should there ever be ‘sales-resistance’? It’s most easily seen in the case of the arms race between the sexes. You might think it’s always a good idea for males and females to get together and coordinate. You’d be wrong, and for an interesting reason. Ultimately because sperms are smaller and more numerous (‘cheaper’) than eggs, females need to be choosier than males. A male is more likely to ‘want’ to mate with a female than the female will ‘want’ to mate with him. Females pay a higher cost if they mate with the wrong male than males pay if they mate with the wrong female. In extreme cases, there is no such thing as the wrong female. Hence males are more likely to escalate salesmanship when trying to persuade females. And females more likely to favour sales-resistance. Where you see high-amplitude signals – bright colours, loud sounds – that means there’s probably sales-resistance. Where there’s no sales-resistance, signals are likely to sink to a conspiratorial whisper. Conspicuous signals are costly, if not in energy, in risk of attracting predators or alerting prey.
I’ve been a bit terse in condensing two full-sized papers into four paragraphs. It should become clearer when I now apply it to birdsong. Birdsong is too loud and conspicuous to be a ‘conspiratorial whisper’, so let’s go for the other extreme: increased sales-resistance fomenting exaggerated efforts to manipulate. Is birdsong an attempt to manipulate the behaviour of females and other males: an attempt to change their behaviour to the advantage of the singer?
If biologists wish to manipulate the behaviour of a bird, what can they do? This chapter has already introduced one possibility that birds themselves, unfortunately for them, cannot do: electrical stimulation of another’s brain through implanted electrodes. The Canadian surgeon Wilder Penfield pioneered the technique on human patients whose brains were undergoing surgery for other reasons. By exploring different parts of the cerebral cortex, he was able to jerk specific muscles into action like a puppeteer pulling strings. When he drew a map of which parts of the brain pulled which muscles, it looked like a caricature of a human body, the so-called ‘motor homunculus’ (there’s also a ‘sensory homunculus’ on the left-hand side of the picture, which looks rather similar). The grotesque exaggeration of the homunculus’s hand goes some way towards explaining the formidable skill of a concert pianist, for example. And the large brain area given over to the lips and tongue is no doubt related to speech. The German biologist Erich von Holst, working with chickens in a deeper part of the brain, the brain stem, was able to control what might be called the bird’s ‘mood’ or ‘motivation’, resulting in changes to the observed behaviour, including ‘guiding hen to nest’ and ‘uttering call warning of predator’. I repeat that these operations are painless, by the way. There are no pain receptor nerves in the brain.
Now, a male nightingale might well ‘wish’ he could implant electrodes in a female’s brain and control her behaviour like a puppet. He can’t do that, he’s no von Holst, and he has no electrodes. But he can sing. Might song have something like the same manipulative effect? No doubt he might benefit, if only he could inject hormones into her bloodstream. Again, he can’t literally do that. But evidence on ring doves and canaries suggests that birds can do something close to it. Male doves vigorously court females with a display called the bow-coo. The bow is a characteristic movement resembling an unusually obsequious human bow, and it is accompanied by an equally characteristic coo, consisting of a staccato note followed by a purring glissando. A week’s exposure to a bow-cooing male reliably causes massive growth of a female’s ovary and oviduct, with accompanying changes in sexual, nest-building, and incubation behaviour. This was shown by the American animal psychologist Daniel S Lehrman. Lehrman went on to show that the behaviour of male ring doves has a direct effect on the hormones circulating in female bloodstreams. Parallel work by Robert Hinde and Elizabeth Steel in Cambridge on nest-building behaviour in female canaries showed the same thing.
The ring dove and canary type experiments have not been done on nightingales, but it probably is generally the case that male birdsong changes the hormonal state of females. Male song manipulates female behaviour, as though the male had the power to inject her with chemicals, presumably nightingales no less than other species.
My heart aches, and a drowsy numbness pains
My sense, as though of hemlock I had drunk,
Or emptied some dull opiate to the drains
One minute past, and Lethe-wards had sunk.
John Keats was not a bird, but his brain was a vertebrate brain like a female nightingale’s. The male nightingale song drugged him – almost to death in his poetic fancy. If it can so intoxicate the mammal Keats, might it not have a yet more powerful effect on the vertebrate brain that it was designed to beguile, the brain of another nightingale? To answer yes, we hardly need the testimony of the dove and canary experiments. I believe natural selection has shaped the male nightingale’s song, perfecting its narcotic power to manipulate the behaviour of a female, presumably by causing her to secrete hormones.
But now, let’s return to learning, and the deafening experiments. The evidence shows that young white-crowned sparrows and song sparrows teach themselves to sing with reference to a template. Young white-crowneds need to hear song in order to make their ‘recording’ of the template. But any old song won’t do. They have to hear the song of their own species. This shows that, even when the template is recorded, there is an innate component to it, built in by the genes. And in the case of the song sparrow, it doesn’t even need to be recorded.
I suggested above that birdsong might be appreciated as music, enjoyed aesthetically by the birds themselves. We are now in a position to spell out the argument. The male teaches himself to sing by comparing his ‘random’ burblings against a template. The template serves as reward, positively reinforcing those random attempts that happen to match it. Reflect, now, that the male songster has a brain much like the female he later hopes to manipulate. When he teaches himself to sing, he is finding out which fragments of song appeal to a bird of his own species (himself … but later, a female). What is that, if not the employment of aesthetic judgment?
Burble. I like it (conforms to my template). Repeat it.
Burble warble. Ooooh, that’s even better. I like that very much.
It really turns me on. Repeat that too. YES!
What turns him on will probably turn a female on too, for they are, after all, members of the same species with the same typical brain of the species. At the end of the developmental period when the final adult song has been perfected, it will be equally beguiling to the singer himself and his female target. He learns to sing whichever phrases turn him on. There seems no powerful reason to deny that both enjoy an aesthetic experience – as did John Keats when he heard the nightingale.
We’ve come a long way from the idea of reward as generalised ‘drive reduction’. And we’ve arrived at what I think is a much more interesting place. The lesson of these experiments on birdsong is that reward can be a highly specific stimulus, or stimulus-complex, ultimately laid down by genes: what Konrad Lorenz, one of the fathers of ethology, dubbed the ‘Innate Schoolmarm’.
If this is right, we should predict the following result in a Skinner Box. A young song sparrow who has never heard song should learn to peck a key that yields the sound of song sparrow, but no other species’ song. That hasn’t been done, but various similar experiments have. Joan Stevenson found that chaffinches preferred to settle on a perch attached to a switch that turned on chaffinch song. However, the control sound for comparison was white noise, not the song of another species. Her chaffinches, moreover, were not naive but wild caught. Her method was adopted by Braaten and Reynolds with hand-reared, naive zebra finches and using starling song for comparison instead of white noise. They showed a clear preference for perches that played zebra finch song rather than starling song. It would be nice to do a big experiment with, say, naive young songbirds of six different species, with six perches, each perch playing one of the six songs. We should predict that each species should learn to sit on the perch that played their own species song. It wouldn’t be an easy experiment. Hand-rearing baby songbirds is hard work. A neat design might be to give each baby to foster parents of one of the other six species.
The template of song sparrows is innate. The ‘recorded’ template of young white-crowned sparrows, laid down early in life before they start singing, looks like the kind of learning called ‘imprinting’, most closely associated with Konrad Lorenz and his pursuing geese. Imprinting was first recognised in nidifugous baby birds.
Nidifugous, from the Latin, means ‘fleeing the nest’. Nidifugous hatchlings start life equipped with warm and protective downy feathers and well-coordinated limbs. Examples are ducklings, goslings, moorhen chicks, chicken chicks, ground-nesting species generally. Within hours of hatching, as soon as their feathers are dry, nidifugous chicks are up and about, walking competently, looking around alertly, pecking at food prospects, and dogging parental footsteps. The opposite of nidifugous is nidicolous. All songbirds are nidicolous. Nidicolous bird species typically nest in trees. The babies are helpless, naked, incapable of walking (they’re in a nest balanced up a tree, where would they walk to?), incapable of feeding themselves but with a huge gaping beak, a begging organ into which their parents tirelessly shovel food. Many seabirds such as gulls are nidifugous in that they hatch with downy feathers and don’t gape for food. But they are dependent on the parents bringing food that they regurgitate for the chicks.
Mammals, too, have their own equivalent to nidifugous (think gambolling lambs; and wildebeest calves must follow the herd on the day they’re born) and nidicolous (baby mice are hairless and helpless). Man is a nidicolous species. Our babies are almost completely helpless. There has been an evolutionary trade-off between a pressure towards a bigger brain, conflicting with the difficulty of being born imposed by a large head. The result was to push our babies towards being born earlier, before the head became insufferably (for the mother) large to push out. The result was to make us even more helplessly nidicolous than other ape species.
Nidifugous species, both mammals and birds, are in danger if they become separated from their parent(s), and this is where imprinting comes in. Nidifugous babies, as soon as they hatch, do something equivalent to taking a mental photograph of the first large moving object they see. They then follow it about, at first very closely, then venturing gradually further away as they grow older. The first moving object they see is usually their parent, so the system works fine in nature. Goslings hatched in an incubator, however, tend to imprint on a human carer, for example Konrad Lorenz.
The idea of imprinting in mammals is imprinted in child minds by the nursery rhyme ‘Mary had a little lamb’ (Everywhere that Mary went / The lamb was sure to go). Imprinted animals, both birds and mammals, often retain their mental photograph into adulthood and attempt to mate with creatures (such as humans) who resemble it. One of the reasons zoos have difficulty with breeding is that the frustrated animals hanker after their keepers.
Imprinting may or may not be a special kind of learning. Some say it’s just a special case of ordinary learning. It’s controversial. Either way, it’s a nice example of a recent, ‘top layer’ palimpsest script. The genes could have equipped the animal with a built-in image or specification of precisely what to follow, what to mate with, what song to sing. Instead, they equip the animal with rules for colouring in the details.
Reinforcement learning and imprinting are not the only kinds of learning by which an animal, during its own lifetime, supplements the inherited ancestral wisdom. Elephants make important use of traditional knowledge. The brains of old matriarchs contain a wealth of knowledge about such vital matters as where water can be found. Young chimpanzees learn from their elders skills such as using a stone as a hammer to crack nuts, and preparing a twig to probe termite nests. The handover from adept to apprentice is a kind of inheritance, but it is memetic, not genetic. This is why these skills are practised in particular local areas and not others. The skill of sweet potato washing in Japanese macaques is another example. So is pecking through the foil or cardboard lids of milk bottles by British tits, in the days when milk was delivered daily on the doorstep. In this case, the skill was seen to radiate geographically outwards from focal points, in the manner of an epidemic.
What else equips animals to improve on their genetic endowment, apart from learning? Perhaps the most important example of a ‘memory’ not mediated by the brain is the immune system. Without it, none of us would have survived our first infection. Immunology is a huge subject, too big for me to do it justice in this book. I’ll say a few words, just enough to make the point that genes don’t attempt the impossible task of equipping bodies with information about all the bacteria, viruses, and other pathogens that they might ever encounter. Instead, genes furnish us with tools for ‘remembering’ past infections, forearming us against future infection. We carry not just the genetic book of the dead (the ancestral past) but a special molecular book in which is written a continually updated medical record of our infections and how we dealt with them.
Geese imprinted on Konrad Lorenz. A special kind of learning, which casts light on the mind of birds
Bacteria, too, suffer from infection – by viruses called bacteriophages, or phages for short – and they have their own immune system, which is rather different from ours. When a bacterium is infected, it stores a copy of part of the viral DNA within its own single circular chromosome. These copies have been called ‘mug shots’ of criminal viruses. Each bacterium sets aside a portion of its circular chromosome as a kind of library of these mug shots. The mug shots will later be used to apprehend criminals in the form of the same or related viruses making a reappearance. The bacterium makes RNA copies of the mug shots. These RNA images of ‘criminal’ DNA are circulated through the interior of the bacterial cell. If a virus of a familiar type should invade, the appropriate mug shot RNA binds to it, and special protein enzymes cut up the joined pair, rendering the virus harmless.
The bacterium needs a way to label the mug shots, so they aren’t confused with its own DNA. They are labelled by the presence of adjacent nonsense sequences of DNA, which are palindromes called CRISPR: Clustered Regularly Interspaced Short Palindromic Repeats. Each time a bacterium is assailed by a new kind of virus, another CRISPR-flanked mug shot is added to the CRISPR region of the chromosome. It’s another story, but CRISPR has become famous because scientists have discovered a way in which the bacterial skill can be borrowed for the human purpose of editing genomes.
The vertebrate immune system works rather differently. It’s more complicated but we too have a ‘memory’ of pathogens of the past. Our immune system is then able to mount a rapid response, should any of those old enemies venture to return. This is why those of us who have had mumps or measles can safely mingle with victims, confident that we shall not get the disease a second time. And the enormous boon of vaccination works by tricking the immune system into building up a false memory, normally by injecting either a killed strain or a weakened strain of the pathogen.
The Covid-19 pandemic was largely stopped in its tracks, saving thousands of lives, by a wonderful new type of vaccine, the mRNA vaccine. The role of mRNA (messenger RNA) is to convey coded messages from DNA in the nucleus to where proteins are made to the code’s specification. Now, here’s how mRNA vaccines work. Instead of injecting a killed or weakened strain of the dangerous virus, a harmless protein in its jacket is first sequenced. The genetic code appropriate to that protein is then written into mRNA. The mRNA does its thing, which is to code the synthesis of protein – in this case the harmless jacket protein of the Covid virus. And then, the immune system does its thing and attacks the virus if it enters the body, recognising it by the protein in its jacket.
What is especially interesting, in pursuit of our analogy between learning and evolution, is that the vertebrate immune system’s ‘memory’ (unlike the bacterial one) works in a kind of Darwinian way, by an internal version of natural selection, within the body. But that is another story, beyond our scope here.
The immune system, and the brain, are the two rich data banks in which entries are written during the animal’s own lifetime, to update the genetic book of the dead, or ‘colour in the details’. More minor examples need mentioning for the sake of completeness. Darkening of the skin is a kind of memory of lying out in the sun. It provides useful screening against the damage that the sun’s rays, especially ultraviolet, can wreak, for example in causing skin cancers. This is a case where genetic and post-genetic scripts both contribute. People whose ancestors have lived many generations in fierce tropical sun tend to be born with dark skin, for example native Australians, many Africans, and people from the south of the Indian sub-continent. By contrast, those whose ancestors have lived many generations at higher latitudes are at risk from too little sun. They tend to lack Vitamin D and hence are prone to rickets. Genetic natural selection at high latitudes has therefore favoured lighter skins. That’s all written in the genetic book of the dead. But this chapter is about palimpsest scripts written after birth, and here is where suntan comes in. Browning in the sun, a post-birth ‘colouring-in’, achieves in light-skinned, high-latitude people a temporary approach towards what is written into the genome of tropical peoples. You could think of the two as short-term memory and long-term memory of sunlight.
Another example is acclimatisation to high altitude. The higher you go, the thinner the atmosphere, where lack of oxygen causes ‘mountain sickness’, whose symptoms include headaches, dizziness, nausea, and complications of pregnancy. People whose ancestors have long lived at high altitude have evolved genetic adaptations such as elevated haemoglobin levels in the blood. Those ‘memories’ of ancestral natural selection are written in the genetic book of the dead. Interestingly, the details differ between Andean and Himalayan peoples, not surprisingly because they have independently, over 10,000 years or more, adapted to a lack of oxygen in mountainous regions widely separated from each other. There are several routes to acclimatisation, and it is not surprising that different mountain peoples have followed different evolutionary paths.
Once again, ancestral scripts can be over-written during the animal’s own lifetime. Lowland people who move to high areas can acclimatise. In 1968, when the Olympic Games were held in Mexico City, national teams deliberately arrived early, in order to train at the high altitude (2,200 metres, more than 7,000 feet) of the Anahuac Plateau. Changes that develop during a period of weeks living at high altitude are written into the post-birth palimpsest layer. As with skin colour, they mimic the older, gene-authored scripts.
Talking of skin colour, the ‘paintings’ of Chapter 2 were all done by ancestral genes, replaying ancestral worlds. But there are some animals who can repaint their skin on the fly, to match the changing background they happen to be sitting on at any given moment. This is another example of the non-genetic book of the living. Chameleons are proverbial, but they aren’t the top virtuosi when it comes to impromptu skin artistry. Flatfish such as plaice can change not just their colour but also their patterning. The one above is capable of changing its colour to match the yellow background on which it now sits. But you only have to take one look at it to read it as a detailed description of the lighter bottom it has just moved off, with its mottled pattern projected by shimmering light from surface ripples.
Even flatfish are upstaged by octopuses and other cephalopod molluscs, who have perfected the art of dynamic cross-dressing to an astonishing extent. And they, uniquely in the animal kingdom, do their changes at high speed. Roger Hanlon, while diving off Grand Cayman, saw a clump of brown seaweed suddenly turn ghostly white and swim rapidly away in a puff of sepia smoke. It was an octopus, with a perfect painting of brown seaweed all over its skin. As Hanlon approached, an emergency order from the octopus brain twitched the muscles controlling the tiny bags of pigment peppering the skin. Instantaneously, the whole surface changed colour from perfect camouflage (trying not to be noticed by predators) to scary white (startling would-be predators). Finally, the puff of dark brown ink deflects the attention of would-be predators away from the fleeing octopus.
Thaumoctopus mimicus
Sea snake
Thaumoctopus mimicus
Flounder
Hanlon saw an octopus (upper right) in Indonesian waters, Thaumoctopus mimicus, who mimicked a flounder (lower right), not just its appearance but also its behaviour, stopping and starting in jerky glides over the sand surface. What’s the point? Hanlon is unsure, but he suspects it deceives predators who like to bite off a tentacle but cannot cope with a substantial flatfish. This octopus also can put on a show with its tentacles (upper left), making each one resemble a venomous sea snake (lower left) common in tropical waters. Cephalopods can even change their skin’s texture, ruffling up or puckering it into extraordinary shapes. A colleague once dramatised their other-world strangeness by beginning a lecture on Cephalopods: ‘These are the Martians.’
The main thesis of this book is that the animal can be read as a description of much older, ancestral environments. This chapter has shown how further details are added, on top of the ancestral palimpsest scripts. Earlier chapters invoked a future scientist, SOF, presented with an animal and challenged to read its body and reconstruct the environments that shaped it. There, we spoke only of ancestral environments, described in the genomic database and its phenotypic manifestations. In this chapter we’ve seen how SOF could supplement her reading of ancestral environments, by additional readings of the more recent past, including the other two great databases that supplement the genes, namely the brain and the immune system. Today’s doctors can read your immune system database and reconstruct a moderately complete history of the infections you have suffered – or been vaccinated against. And if SOF could read what is written in the brain (a big if, she really would have to be a scientist of the future), she could reconstruct much detail of the animal’s past environments in its own lifetime.
Experience, either literal experience stored in the brain as memories, disease experience, or genetic ‘experience’ sculpted into the genome by natural selection, enables an animal to predict (behave as if predicting) what will happen next. But there’s one more trick that the brain can pull off in order to foretell the future: simulation, or imagination. Human imagination is a much grander affair than this but, from the point of view of an animal’s survival, and our analogy between natural selection and learning, we could regard imagination as a kind of ‘vicarious trial and error’. Unfortunately, that particular phrase has been usurped by rat psychologists. A rat in a ‘maze’ (usually just a choice between turning left or right) will sometimes physically vacillate, looking left, right, left, right before finally making up its mind. This ‘VTE’ may be a special case of imagining alternative futures, but it’s probably safest if I reluctantly surrender the phrase itself to the rat-runners and not use it here. Instead, I’ll prefer an analogy with computer simulation: the animal’s brain simulates likely consequences of alternative actions internally, thereby sparing itself the dangers of trying them out externally in the real world.
I said the human imagination is a much grander affair. It finds expression in art and literature. Words written by one person can call up an imagined scene in the brain of another. Gertrude’s lament for Ophelia can move a reader to tears four centuries after the poet’s death. Less ambitiously, let me ask you to imagine a baboon atop a steep cliff. Someone has balanced a plank over the edge of the cliff. Resting at the far end of the plank, over the abyss, is a bunch of bananas. Imagine them, yellow and tempting. The baboon is indeed tempted to venture out along the plank. However, his brain internally simulates the consequence, sees that his extra weight would topple the plank – imagines himself tumbling to his death. So he refrains.
Let’s now imagine a range of brains faced with the banana on the plank. First, the genetic book of the dead can build in an innate fear of heights. I myself experience a tingling of the spine, which inhibits me from walking within a metre of the edge of a precipice such as the Cliffs of Moher in Western Ireland. This, even when there’s no wind and no reason to suppose that I would fall.
The visual cliff
A whole genre of experimentation, the so-called ‘visual cliff’ experiment, has been devised to investigate fear of heights. The baby in the picture is quite safe: there’s strong glass over the ‘cliff’. I recently visited one of the world’s tallest buildings where one could stand on toughened glass looking down on the street far below. Perfectly safe, and I watched others walk on the glass, but I avoided doing so myself. Irrational, but innate fears are hard to conquer. Perhaps an innate fear of heights is inherited from tree-climbing ancestors who survived because they possessed it. Not everyone succumbs, of course. These New York construction workers are enjoying a relaxed lunch with evident (though incomprehensible to me) nonchalance.
Death by falling is the crudest route through which a fear of heights might be built into animals. Another way is by learning, reinforced by pain. Young baboons who fall down smaller cliffs are not killed, but they experience pain. Pain, as we’ve seen, is a warning: ‘Don’t do that again. Next time the cliff might be higher, and it will kill you.’ Pain is a kind of vicarious, relatively safe substitute for death. Pain stands in for death in the analogy between learning and natural selection.
The ‘detour problem’
But now, since you are human with a human power of imagination, you are probably simulating in your brain an unusually bright baboon. He sees himself, in his own imagination, pulling the plank carefully inwards, complete with bananas. Or reaching out with a stick and nudging the bananas along the plank towards him. Probably only highly evolved brains are capable of such simulations. Even dogs (above) perform surprisingly poorly on the so-called ‘detour problem’. But if he succeeds, this imaginative baboon risks no pain and doesn’t fall to his death but does it all by internal simulation. He simulates the fall in his imagination, and consequently refrains from venturing out along the plank. He then simulates the safe solution to the problem and gets the bananas.
I need hardly say that internal simulation of dangerous futures is preferable to the actual actions. Provided, of course, that the simulation leads to accurate prediction. Aircraft designers find it cheaper and safer to test model wings in wind tunnels rather than actual wings on real aeroplanes. And even wind tunnel models are more expensive than computer simulations or analytical calculations, if these can be done. Simulation still leaves some room for uncertainty. The maiden flight of a new plane is still an informative event, however rigorously its parts have been subjected to ordeal by wind tunnel or computer simulation.
Once a sufficiently elaborate simulation apparatus is in place in a brain, emergent properties spring up. The brain that can imagine how alternative futures might affect survival can also, in the skull of a Dante or a Hieronymus Bosch, imagine the torments of Hell. The neurons of a Dalí or an Escher simulate disturbing images that will never be seen in reality. Non-existent characters come alive in the head of the great novelist and in those of her readers. Albert Einstein, in imagination, rode a sunbeam to his place among the immortals with Newton and Galileo. Philosophers imagine impossible experiments – the brain in a vat (‘Where am I?’), atom-for-atom duplication of a human (which ‘twin’ would claim the ‘personhood’?). Beethoven imagined, and wrote down, glories that he tragically could never hear. The poet Swinburne happened upon a forsaken garden on a sea cliff, and his imagination revived a pair of long-dead lovers whose eyes went seaward, ‘a hundred sleeping years ago’. Keats reconstructed the ‘wild surmise’ with which stout Cortez and all his men stared at the Pacific, ‘silent upon a peak in Darien’.
The ability to perform such feats of imagination sprang, emergently, from the Darwinian gift of vicarious internal simulation within the safe confines of the skull, of predicted alternative actions in the unsafe real world outside. The capacity to imagine, like the capacity to learn by trial and error, is ultimately steered by genes, by naturally selected DNA information, the genetic book of the dead.
8 The Immortal Gene
The central idea of The Genetic Book of the Dead grows out of a view of life that may be called the gene’s-eye view. It has become the working assumption of most field zoologists studying animal behaviour and behavioural ecology in the wild, but it has not escaped criticism and misunderstanding, and I need to summarise it here because it is central to the book.
There are times when an argument can helpfully be expressed by contrast with its opposite. Disagreement that is clearly stated deserves a clear reply. I could hypothetically invent the opposite of the gene’s-eye view, but fortunately I don’t need to because the diametric opposite has been put, articulately and clearly, by my Oxford colleague (and incidentally my doctoral examiner, on a very different subject long ago) Professor Denis Noble. His vision of biology is alluring, and is shared by others whose expression of it is less explicit and less clear. Noble is clear. He ringingly hits a nail on the head, but it’s the wrong nail. Here is his lucid and unequivocal statement, right at the beginning of his book Dance to the Tune of Life:
This book will show you that there are no genes ‘for’ anything. Living organisms have functions which use genes to make the molecules they need. Genes are used. They are not active causes.
That is precisely and diametrically wrong, and it will be my business in this chapter to show it.
If genes are not active causes in evolution, almost all scientists now working in the fields known as Behavioural Ecology, Ethology, Sociobiology, and Evolutionary Psychology have been barking up a forest of wrong trees for half a century. But no! ‘Active causes’ is precisely what genes must be: necessarily so if evolution by natural selection is to occur. And, far from being used by organisms, genes use organisms. They use them as temporary vehicles, which they exploit in the service of journeying to future generations. This is not a trivial disagreement, no mere word game. It is fundamental. It matters.
A physiologist of distinction, Denis Noble is captivated by the shattering complexity of the organism, of every last one of its trillions of cells. He sets out to impress his readers with the intricate co-dependency of all aspects of the living organism. As far as this reader is concerned, he succeeds. He sees every part as working inextricably with every other part in the service of the whole. In that service – and this is where he goes wrong – he sees the DNA in the nucleus of a cell as a useful library to be drawn upon when the cell needs to make a particular protein. Go into the nucleus, consult the DNA library there, take down the manual for making the useful protein, and press it into service. I devised that characterisation of Noble’s position during a public debate with him in Hay-on-Wye, and he vigorously nodded his assent. DNA, in Noble’s view, is the servant of the organism, in just the same way as the heart or the liver or any cell therein. DNA is useful to make a particular enzyme when you need it, just as the enzyme is useful for speeding up a chemical reaction … and so on.
Dance to the Tune of Life has the subtitle ‘Biological Relativity’. Noble’s usage of ‘relativity’ has only a tenuous and contrived connection with Einstein’s, but it exactly matches that of the historian Charles Singer in A Short History of Biology:
The doctrine of the relativity of functions is as true for the gene as it is for any of the organs of the body. They exist and function only in relation to other organs.
Now here is Noble some ninety years later. He has the advantage over Singer in that we now know genes are DNA. But his sentiment about biological relativity, in conjunction with the quotation above, resonates perfectly with Singer’s.
The principle of Biological Relativity is simply that there is no privileged level of causation in biology.
I shall argue that, no matter how complicatedly interdependent the parts of a living organism are when we are talking physiology, when we move to the special topic of evolution by Darwinian natural selection there is one privileged level of causation. It is the level of the gene. To justify that is the main purpose of this chapter.
Here’s Singer’s whole vitalistic passage from which I took the above quotation. It’s the peroration of his book and is a perfect prefiguring of Noble’s ‘relativity’.
Further, despite interpretations to the contrary, the theory of the gene is not a ‘mechanist’ theory. The gene is no more comprehensible as a chemical or physical entity than is the cell or, for that matter, the organism itself. Further, though the theory speaks in terms of genes as the atomic theory speaks in terms of atoms, it must be remembered that there is a fundamental distinction between the two theories. Atoms exist independently, and their properties as such can be examined. They can even be isolated. Though we cannot see them, we can deal with them under various conditions and in various combinations. We can deal with them individually. Not so the gene. It exists only as a part of the chromosome, and the chromosome only as part of a cell. If I ask for a living chromosome, that is, for the only effective kind of chromosome, no one can give it to me except in its living surroundings any more than he can give me a living arm or leg. The doctrine of the relativity of functions is as true for the gene as it is for any of the organs of the body. They exist and function only in relation to other organs. Thus the last of the biological theories leaves us where the first started, in the presence of a power called life or psyche which is not only of its own kind but unique in each and all of its exhibitions.
Watson and Crick blew that out of the water in 1953. The triumphant field of digital genomics that they initiated falsifies every single one of Singer’s sentences about the gene. It is true but trivial that a gene is impotent in the absence of its natural milieu of cellular chemistry. Here’s Noble again, bringing Singer up to date but agreeing with his sentiment:
There really is nothing alive in the DNA molecule alone. If I could completely isolate a whole genome, put it in a petri dish with as many nutrients as we may wish, I could keep it for 10,000 years and it would do absolutely nothing other than to slowly degrade.
Obviously a gene in a petri dish cannot do anything, and it would degrade as a physical molecule within months, let alone 10,000 years. But the information in DNA is potentially immortal, and causally potent. And that is the whole point. Never mind the physical molecule and never mind the petri dish. Let the sequence of A, T, C, G triplet codons of an organism’s genome be written on a long paper scroll. Or, no, paper is too friable. To last 10,000 years, carve the letters deep in the hardest granite. To be sure, world-spanning ranges of highland massif would still be too small, but that is a superficial difficulty. In 10,000 years, if scientists still walk the Earth, they will read the sequence and type it into a DNA-synthesising machine such as we already have in early form. They’ll have the embryological knowhow to create a clone of whoever donated the genome in the first place (just a version of the way Dolly the sheep was made). Of course, the DNA information would need the biochemical infrastructure of an egg cell in a womb, but that could be provided by any willing woman. The baby she bears, an identical twin of its 10,000-year dead predecessor, would be living repudiation of Singer and Noble.
That the information necessary to create the twin could be carved in lifeless granite and left for 10,000 years is a truth that fills me with amazement still, even seventy years after Watson and Crick prepared us for it. Charles Singer would be forced to recant his vitalism, while Charles Darwin, I suspect, would exult.
The point is that, transitory though physical DNA molecules themselves may be, the information enshrined in the nucleotide sequence is potentially eternal. Essential though the surrounding machinery is – messenger RNA, ribosomes, enzymes, uterus and all – they can be provided anew by any woman. But the information in an individual’s DNA is unique, irreplaceable, and potentially immortal. Carving it in granite is a way to dramatise this. But it’s not the practical way. In the normal course of events, DNA information achieves its immortality through being copied. And copied. And copied. Copied indefinitely, potentially eternally, down the generations. Of course, DNA can’t copy itself on its own. Obviously, just as a computer disc can’t copy itself without supporting hardware, DNA needs an elaborate infrastructure of cellular chemistry. But of all the molecules that are involved in the process, however essential they may be for the copying process, only DNA is actually copied. Nothing else in the body is so honoured. Only the information written in DNA.
You might think every part of the body is replicated. Does not every individual have arms and kidneys, and are these not renewed in every generation? Yes, but you’d be utterly wrong if you called it replication in the sense that genes are replicated. Arms and kidneys don’t replicate to make new arms and kidneys. Here’s the acid test, and it really matters. Make a change to an arm, say by a fracture or by pumping iron, and the change is not propagated to the next generation. Make a change in a germline gene, on the other hand, and the mutation may long outlast 10,000 years, copied again and again down the generations.
Before the invention of printing, biblical scriptures were painstakingly copied by scribes at regular intervals to forestall decay. The papyrus might crumble but the information lived on. Scrolls don’t replicate themselves. They need scribes, and scribes are complicated, just as the enzymes involved in DNA replication are complicated. Through the mediation of scribes/enzymes information in scrolls/DNA is copied with high fidelity. Actually, scribes might copy with lower fidelity than DNA replication can achieve. With the best will in the world human copyists make errors, and some zealous scribes were not above a little well-meant improvement. Older manuscripts of Mark 9, 29 quote Jesus as saying that a particular kind of demonic possession can be cured only by prayer. Later versions of the text, not content with mere prayer, say ‘prayer and fasting’. It seems that some zealous scribe, perhaps belonging to a monkish order that especially valued fasting, thought to himself that Jesus must surely have meant to mention fasting, how could he not? So it was scarcely taking a liberty to put the words into his mouth. DNA is capable of higher fidelity of replication than that, but even DNA is not perfect. It does make mistakes – mutations. And in one important respect, DNA is unlike the over-zealous scribe: mutation is never biased towards improvement. Mutation has no way to judge in which direction improvement lies. Improvement is judged retrospectively. By natural selection.
So the information in DNA is potentially eternal even though the physical medium of DNA is finite. And let me repeat why this matters. Only the information contained in DNA is destined to outlive the body. Outlive in a very big way. Most animals die in a matter of years if not months or weeks. Few survive the ravages of decades, almost none centuries. And their physical DNA molecules die with them. But the information in the DNA can last indefinitely. I once attended an evolution conference in America where, at the farewell dinner, we were all challenged to produce an appropriate poem. My limerick ran as follows:
An itinerant Selfish Gene
Said ‘Bodies a-plenty I’ve seen.
You think you’re so clever
But I’ll live for ever:
You’re just a survival machine.’
And I raided Rudyard Kipling for the body’s reply:
What is a body that first you take her,
Grow her up and then forsake her,
To go with the old Blind Watchmaker.
I have emphasised the immortality of the gene in the form of copies. But how big is the unit that enjoys such immortality? Not the whole chromosome: it is far from immortal. With minor exceptions such as the Y-chromosome, our chromosomes don’t march intact down the centuries. They are sundered in every generation by the process of crossing over. For the purposes of this argument, the length of chromosome that should be considered significant in the long run depends upon how many generations it is allowed, by crossing over, to remain intact, when measured against the relevant selection pressures. I expressed this only slightly facetiously in my first book, The Selfish Gene, by saying that the title strictly should have been The slightly selfish big bit of chromosome and the even more selfish little bit of chromosome. A small fragment of chromosome, such as a gene responsible for programming one protein chain, can last 10,000 years. In the form of copies. But only fragments that are successful in negotiating the obstacle course that is natural selection actually do that. It’s arguable that a better book title would have been The Immortal Gene, and I have adopted it as the title of this chapter. As we shall see in Chapter 12, it is no paradox that The Cooperative Gene would also have been appropriate.
How does a gene earn ‘immortality’? In the form of copies, it influences a long succession of bodies so that they survive and reproduce, thereby handing the successful gene on to the next generation and potentially the distant future. Unsuccessful genes tend to disappear from the population, because the bodies they successively inhabit fail to survive into the next generation, fail to reproduce. Successful genes are those with a statistical tendency to inhabit bodies that are good at surviving and reproducing. And they enjoy that statistical tendency, positive or negative, by virtue of the causal influence they exert over bodies. So, we have arrived at the reason why it was profoundly wrong to say that genes are not active causes. Active causes is precisely and indispensably what they must be. If they were not, there could be no natural selection and no adaptive evolution.
‘Cause’ has a testable meaning. How do we ever identify a causal agent in practice? We do it by experimental intervention. Experimental intervention is necessary, because correlation does not imply causation. We remove, or otherwise manipulate, the putative cause, and we strictly must do so at random, a large number of times. Then we look to see whether there tends to be a statistically significant change in the putative effect. To take an absurd example, suppose we notice that the church clock in the village of Runton Acorn reliably chimes immediately after that of Runton Parva. If we’re very naive, we jump to the conclusion that the earlier chiming causes the later. But of course it’s not good enough to observe a correlation. The only way to demonstrate causation is to climb up the church tower in Runton Parva and manipulate the clock. Ideally, we force it to chime at random moments, and we repeat the experiment many times. If the correlation with the Runton Acorn chiming is maintained, we have demonstrated a causal link. The important point is that causation is demonstrated only if we manipulate the putative cause, repeatedly and at random. Of course, nobody would be silly enough to actually do this particular experiment with the church clocks. The result is too obvious. I use it only to clarify the meaning of ‘cause’.
Now back to Denis Noble’s statement that ‘Genes are used. They are not active causes.’ By our ‘church clock’ definition, genes most definitely are active causes because, if a gene mutates (a random change), we consistently observe a change in the body of the next generation – and subsequent generations for the indefinite future. Mutation is equivalent to climbing the Runton Parva tower and changing the clock. By contrast, if there is a non-genetic change in the body (a scar, a lost leg, circumcision, an exaggeratedly muscular arm due to exercise, a suntan, acquired fluency in Esperanto or virtuosity on the bassoon), we do not observe the same thing in the next generation. There is no causal link.
Genetic information, then, is potentially immortal, is causal, and there’s a telling difference between potentially immortal genes that succeed in being actually immortal and potentially immortal genes that fail. The reason some succeed and others fail is precisely that they have a causal influence, albeit a statistical one, on the survival and reproductive prospects of the many bodies that they inhabit, through successive generations and across many bodies through populations. It’s important to stress ‘statistical’. One copy of a good gene may fail to survive to the next generation because the body it inhabits is struck by lightning or otherwise suffers bad luck. More relevantly, one copy of a good gene may happen to find itself sharing a body with bad genes, and is dragged down with them. Statistics enter in because sexual recombination sees to it that good genes don’t consistently share bodies with bad genes. If a gene is consistently found in bodies that are bad at surviving, we draw the statistical conclusion that it is a bad gene. After 10,000 years of recombining, shuffling, recombining again, a gene that remains in the gene pool is a gene that is good at building bodies: in collaboration with the other genes that it tends to share bodies with, and that means the other genes in the gene pool of the species (you may remember from Chapter 1 that the species can be seen as an averaging computer).
In The Selfish Gene, I used the image of the Oxford vs Cambridge Boat Race, the parable of the rowers. Eight oarsmen and a cox all have their part to play, and the success of the whole boat depends upon their cooperation. They must not only be strong rowers, they must be good cooperators, good at melding with the rest of the crew. The rowers, of course, represent genes, and they are arrayed along the length of the boat, as genes are arrayed along a chromosome. It’s hard to separate the roles of the individual oarsmen, so intimate is their cooperation, and so vital is cooperative pulling together for the success of the whole boat. The coach swaps individual rowers in and out of his trial crews. Although it’s hard to judge individual performance by watching them, he notices that certain individuals consistently seem to be members of the fastest trial crews. Other individuals consistently are seen to be members of slower crews. Although single individuals never row on their own, in the long run the best rowers show their mettle in the performance of the successive boats in which they sit.
Natural selection sorts out the good genes from the bad, precisely because of the causal influence of genes on bodies. The practical details vary from species to species. Genes that make for good swimmers are ‘good genes’ in a dolphin gene pool but not in a mole gene pool. Genes that make for good diggers are ‘good genes’ in a mole, wombat, or aardvark gene pool but not in a dolphin or salmon gene pool. Genes for expert climbing flourish in a monkey, squirrel, or chameleon gene pool but not in a swordfish, rhinoceros, or earthworm gene pool. Genes for aerodynamic proficiency flourish in a swallow or bat gene pool though not in a hippo or alligator gene pool.
But however varied the details of ‘good’ and ‘bad’ may be from species to species, the central point remains. Depending on their causal influence on bodies, genes either survive or don’t survive to the next generation, and the next, and the next … ad infinitum. Let me put it more forcefully: any Darwinian process, anywhere in the universe – and I’m pretty sure if there’s life elsewhere in the universe it will be Darwinian life – any Darwinian process depends on trans-generational replicated information, and that information must have a causal influence on its probability of being replicated from one generation to the next. It happens that on our planet the replicated information, the causal agent in the Darwinian process, is DNA. It is wrong, utterly, blindingly, flat-footedly, downright wrong, to deny its fundamental role as a cause in the evolutionary process.
Have I labored the point excessively? Would that it were excessive, but unfortunately there is reason to think that views such as those I have criticized here have been widely influential. Stephen Jay Gould (whose errors were consistently masked by the graceful eloquence with which he expressed them) went so far as to reduce the role of genes in evolution to mere ‘bookkeeping’. The metaphor of the bookkeeper has a dramatic appeal so seductive that it evidently seduced Gould himself. But it’s as wide of the mark as it is possible to be. It is the bookkeeper’s role to keep a passive record of transactions after they happen. When the bookkeeper makes an entry in his ledger, the entry does not cause a subsequent monetary transaction. It is the other way around.
I hope the preceding pages have convinced you that ‘bookkeeping’ is worse than a hollow travesty of the central causal role that genes play in evolution. It is the exact opposite of the truth, a metaphor as deeply wrong as it is superficially persuasive. Gould was also a proponent of ‘multi-level selection’, and this is another respect in which he is seen as an opponent of the gene’s-eye view of evolution (see, for instance, the philosopher Kim Sterelny’s perceptive book Dawkins Versus Gould: Survival of the Fittest). Gould, and others, insisted that natural selection occurs at many levels in the hierarchy of life: species, group, individual, gene. The first thing to say about this is that although there is a persuasive hierarchy, a real ladder, the gene doesn’t belong on it. Far from being the bottom rung of a ladder, far from being on the ladder at all, the gene is set off to one side. Precisely because of its privileged role as a causal agent in evolution. The gene is a replicator. All other rungs in the ladder are vehicles, a term that I shall explain later in this chapter.
As for higher levels of selection, there is, to be sure, a sense in which some species survive at the expense of others. This can look a bit like natural selection at the species level. The native red squirrel in Britain is steadily going extinct as a direct result of the lamentable whim of the 11th Duke of Bedford in the nineteenth century to introduce American grey squirrels. The greys out-compete the smaller reds, and also infect them with squirrel pox, to which they themselves have evolved resistance over many generations in America. Ecological replacement of a species by a competitor species looks superficially like natural selection. But the resemblance is empty and misleading. This kind of ‘selection’ does not foster evolutionary adaptation. It’s not natural selection in the Darwinian sense. You would not say that any aspect of the grey squirrel’s body or behaviour was a device to drive red squirrels extinct, whereas you might happily talk about the Darwinian function of its bushy tail, meaning those aspects of the tail that assisted ancestral squirrels to out-compete rival squirrel individuals of the same species, with a slightly different tail.
In 1988, I published a paper called ‘The Evolution of Evolvability’. This is the closest I have come to supporting something like ‘multi-level selection’. My thesis was that certain body plans, for example the segmented body plans of arthropods, annelids, and vertebrates, are more ‘evolvable’ than others. I quote from that paper:
I suspect that the first segmented animal was not a dramatically successful individual. It was a freak, with a double (or multiple) body where its parents had a single body. Its parents’ single body plan was at least fairly well‑adapted to the species’ way of life, otherwise they would not have been parents. It is not, on the face of it, likely that a double body would have been better adapted … What is important about the first segmented animal is that its descendant lineages were champion evolvers. They radiated, speciated, gave rise to whole new phyla. Whether or not segmentation was a beneficial adaptation during the individual lifetime of the first segmented animal, segmentation represented a change in embryology that was pregnant with evolutionary potential.
I envisioned that my concept of ‘evolvability’ should be regarded as a property of embryology. Thus, a segmented embryology has high evolvability potential, meaning an embryology that lends itself to rich evolutionary divergence. The world tends to become populated by clades with high evolvability potential. A clade is a branch of the tree of life, meaning a group plus its shared ancestor. ‘Birds’ constitutes a clade, for all birds have a single common ancestor not shared by any non-birds. ‘Fish’ is not a clade, because the common ancestor of all fish is shared by all terrestrial vertebrates including us, who are not fish. ‘Mammals’ is a clade, but only if you include so-called ‘mammal-like reptiles’. It would be unhelpful and confusing to call the evolution of evolvability group selection. ‘Clade selection’, a coining of George C Williams, fits the bill.
What other criticisms of the gene’s-eye view should we consider? Many would-be critics have pointed out that there is no simple one-to-one mapping between a gene and a ‘bit’ of body. Though true, that’s not a valid criticism at all, but I need to explain it because some people think it is. You know those gruesome butchers’ diagrams, where a map of a cow’s body is defaced by lines representing named ‘cuts’ of meat: ‘rump’, ‘brisket’, ‘sirloin’, etc? Well, you can’t draw a map like that for domains of genes. There’s no ‘border’ you can draw on the body, marking where the ‘territory’ of one gene ends and that of the next one begins. Genes don’t map onto bits of body; they map onto timed embryological processes. Genes influence embryonic development, and a change in a gene (mutation) maps onto a change in a body. When geneticists notice a gene’s effects, all they are really seeing is a difference between individuals that have one version (‘allele’) of the gene and individuals that don’t. The units of phenotype that geneticists count, or trace through pedigrees, traits such as the Hapsburg jaw, albinism, haemophilia, or the ability to smell freesias, loop the tongue, or disperse the froth on contact with beer, are all identified as differences between individuals. For, of course, countless genes are involved in the development of any jaw, Hapsburg or not; any tongue, loopy or not. The Hapsburg jaw gene is no more than a gene for a difference between some individuals and other individuals. Such is the true meaning whenever anyone talks of a gene ‘for’ anything. Genes are ‘for’ individual differences. And, just as the eyes of a geneticist are focused on individual differences in phenotype, so also, precisely and acutely, are the eyes of natural selection: differences between those who have what it takes to survive and those who don’t.
As for the all-important interactions between genes in influencing phenotype, here’s a better metaphor than the butcher’s map. A large sheet hangs from the ceiling, suspended from hooks by hundreds of strings attached to different places all over the sheet. It may help the analogy to consider the strings as elastic. The strings don’t hang vertically and independently. Instead, they can run diagonally or in any direction, and they interfere with other strings by cross-links rather than necessarily going straight to the sheet itself. The sheet takes on a bumpy shape, because of the interacting tensions in the tangled cat’s-cradle of hundreds of strings. As you’ve guessed, the shape of the sheet represents the phenotype, the body of the animal. The genes are represented by the tensions in the strings at the hooks in the ceiling. A mutation is either a tug towards the hook or a release, perhaps even a severing of the string at the hook. And, of course, the point of the parable is that a mutation at any one hook affects the whole balance of tensions across the tangle of strings. Alter the tension at any one hook, and the shape of the whole sheet shifts. In keeping with the sheet model, many, if not most, genes have ‘pleiotropic’ (multiple) effects, as defined in Chapter 4.
A balance of tensions
For practical reasons, geneticists like to study the minority of genes that do have definable, seemingly singular effects, like Gregor Mendel’s smooth or wrinkled peas, for example. But even such ‘major genes’ often have a surprisingly miscellaneous collection of other pleiotropic effects, sprinkled seemingly at haphazard around the body. And it’s not surprising that this should be so: genes exert their effects at many stages of embryonic development. It’s only to be expected, therefore, that they’ll have pleiotropic consequences even at opposite ends of the body. A change in tension at one hook leads to a comprehensive shapeshift, all over the whole sheet.
There’s no one-to-one mapping, then, from single gene to single ‘bit’ of body. We have no butcher’s map here. But not by a jot or even a tittle does this fact threaten the gene’s-eye view of evolution. However pleiotropic, however complicated and interactive the effects of a gene may be, you can still add them all up to derive a net positive or net negative effect of a change (mutation) in its influence on the body: a net effect on its chances of surviving into the next generation. Such causal influences on a gene’s own survival in the gene pool come unscathed through the complications, notwithstanding numerous interactions with other genes – the other genes with which it jointly affects the tensions in all the strings. When the gene in question mutates, the whole shape of the sheet may shift, with perhaps lots of pleiotropic changes all over the body. But the net effect of all these changes, in different parts of the body, and in interaction with many other genes, must be either positive or negative (or neutral) with respect to survival and reproduction. That is natural selection.
The tension in the genetic strings is affected too by environmental influences. See these as yet more strings tugging from the side, rather than from hooks in the ceiling. The developing animal is, of course, influenced by the environment as well as by the genes, always in interaction with the genes. But again, this doesn’t matter one iota to the gene’s-eye view of evolution. To the extent that, under available environmental conditions, a change in a gene causes a change in that gene’s chances of making it through the generations (either positive or negative), natural selection will occur. And natural selection is what the gene’s-eye view is all about.
So much for that criticism of the gene’s-eye view. What else do we have? Granted that genes are active causes in evolution, it is the whole individual body that we observe behaving as an active agent. This fact, too, is often wrongly seen as a weakness of the gene’s-eye view. Yes, of course, it is the whole animal who possesses executive instruments with which to interact with the world – legs, hands, sense organs. It’s the whole animal who restlessly searches for food, trying first this avenue of hope, then switching to another, showing all the symptoms of questing appetite until consummation is reached. It is the individual animal who shows fear of predators, looks vigilantly up and around, jumps when startled, runs in evident terror when pursued. It is the individual animal who behaves as a unitary agent when courting the opposite sex. It is the individual animal who skilfully builds a nest, and works herself almost to death caring for her young.
The animal, the individual animal, the whole animal, is indeed an agent, striving towards a purpose, or set of purposes. Sometimes the purpose seems to be individual survival. Often it is reproduction and the survival of the individual’s children. Sometimes, especially in the social insects, it is the survival and reproduction of relatives other than children – sisters and nieces, nephews and brothers. My late colleague WD Hamilton (he of the palimpsest postcard in Chapter 1) formulated the general definition of the exact mathematical quantity that an individual under natural selection is expected to maximise as it engages in its purposeful striving. It includes individual survival. It includes reproduction. But it includes more, because genes are shared with collateral relatives, and gene survival can therefore be fostered by enabling the survival and reproduction of a sister or a nephew. He gave a name to the exact quantity that an individual organism should strive to maximise: ‘inclusive fitness’. He condensed his difficult mathematics into a long and rather complicated verbal definition:
Inclusive fitness may be imagined as the personal fitness which an individual actually expresses in its production of adult offspring as it becomes after it has been first stripped and then augmented in certain ways. It is stripped of all components which can be considered as due to the individual’s social environment, leaving the fitness which he would express if not exposed to any of the harms or benefits of that environment. This quantity is then augmented by certain fractions of the quantities of harm and benefit which the individual himself causes to the fitnesses of his neighbours. The fractions in question are simply the coefficients of relationship appropriate to the neighbours whom he affects: unity for clonal individuals, one-half for sibs, one-quarter for half sibs, one-eighth for cousins … and finally zero for all neighbours whose relationship can be considered negligibly small.
Pretty convoluted? A bit hard to read? Well, it has to be convoluted because inclusive fitness is a hard idea. It’s necessarily convoluted in my view because looking at it from the individual’s point of view is an unnecessarily convoluted way of thinking about Darwinism. It all becomes blessedly simple if you dispense with the individual organism altogether and go straight to the level of the gene. Bill Hamilton himself did this in practice. In one of his papers, he wrote:
let us try to make the argument more vivid by attributing to the genes, temporarily, intelligence and a certain freedom of choice. Imagine that a gene is considering the problem of increasing the numbers of its replicas, and imagine that it can choose between causing purely self-interested behaviour by its bearer … and causing ‘disinterested’ behaviour that benefits in some way a relative.
See how clear and easy to follow that is, compared to the previous quotation on inclusive fitness. The difference is that the clear passage adopts the gene’s-eye view of natural selection. The difficult passage is what you get when you re-express the same idea from the point of view of the individual organism. Hamilton gave his blessing to my half-humorous informal definition: ‘Inclusive fitness is that quantity that an individual will appear to be maximising, when what is really being maximised is gene survival.’
Role
Maximises
Gene
Replicator
Survival
Organism
Vehicle
Inclusive fitness
Bill Hamilton
The individual organism, in my terminology, is a ‘vehicle’ for survival of copies of the ‘replicators’ that ride inside it. The philosopher David Hull got the point after an extensive correspondence with my then student Mark Ridley, but he substituted the word ‘interactor’ for my ‘vehicle’. I never quite understood why. Depending on your preference you can see either the vehicle or the replicator as the agent that maximises some quantity. If it’s the vehicle, then the quantity maximised is inclusive fitness, and rather complicated. But equivalently, if it’s the replicator, the quantity maximised is simple: survival. I don’t want to downplay the importance of vehicles as units of action. It is the individual organism who possesses a brain to take decisions, based on information supplied by senses, and executed by muscles. The organism (‘vehicle’) is the unit of action. But the gene (‘replicator’) is the unit that survives. On the gene’s-eye view, the very existence of vehicles should not be taken for granted but needs explaining in its own right. I essayed a kind of explanation in ‘Rediscovering the Organism’, the final chapter of The Extended Phenotype.
Replicators (on our planet, stretches of DNA) and vehicles (on our planet, individual bodies) are equally important entities, equally important but they play different, complementary roles. Replicators may once have floated free in the sea but, to quote The Selfish Gene, ‘they gave up that cavalier freedom long ago. Now they swarm in huge colonies, safe inside gigantic lumbering robots’ (individual bodies, vehicles). The gene’s-eye view of evolution does not play down the role of the individual body. It just insists that that role (‘vehicle’) is a different kind of role from that of the gene (‘replicator’).
Successful genes, then, survive in bodies down the generations, and they cause (in a statistical sense) their own survival by their ‘phenotypic’ effects on the bodies that they inhabit. But I went on to amplify the gene’s-eye view by introducing the notion of the extended phenotype. For the causal arrow doesn’t stop at the body wall. Any causal effect on the world at large – any causal effect that can be attributed to the presence of a gene as opposed to its absence, and that influences the gene’s chances of survival, may be regarded as a phenotypic effect, of Darwinian significance. It has only to exert some kind of statistical influence on the chances, positive or negative, on that gene’s surviving in the gene pool. I must now revisit the extended phenotype, for it is, to me, an important part of the gene’s-eye view of evolution.
Alternative titles for The Selfish Gene, all true to its content
9 Out Beyond the Body Wall
Imagine the furore if Jane Goodall reported seeing chimpanzees building an amazing stone tower in a forest clearing. They meticulously select stones of the correct shape for the purpose, rotating each one until it snugly fits neighbouring stones. Then the chimps cement it securely in place before picking out another stone. They evidently like to use two radically different sizes of stones, small ones to build the walls themselves, and much larger ones to provide outer fortification and structural strength, the all-important supporting walls. The discovery would be a sensation, headline news, the subject of breathless BBC discussions. Philosophers would jump on it, there’d be passionate debates about personhood, moral rights, and other topics of philosophical moment. The tower is ill-suited to housing its builders. If not functional, then, is it some kind of monument? Does it have ritual or ceremonial significance like Stonehenge? Does the tower show that religion is older than mankind? Does it threaten the uniqueness of man?
The edifice pictured is a real animal construction, but not one built by chimpanzees; the reality is much smaller, and it doesn’t stand up like a monument but lies flat on the bottom of a stream. It is the house of a little insect, the larva of a caddis fly, Silo pallipes. Caddis adults fly in search of mates and live only a few weeks, but their larvae grow for up to two years under water, living in mobile homes that they build for themselves out of materials gathered from their surroundings, cementing them with silk that they secrete from glands in the head. In the case of Silo pallipes (see top left of picture) the building material is local stone. Its astonishing building skills were unravelled by Michael Hansell, now our leading expert on animal architecture in general.
These larvae are master masons. Just look at the delicate placing of the small stones between the carefully chosen large ones buttressing the sides. Hansell showed how they select stones, choosing by size and shape but not by weight. Ingenious experiments in whichhe removed parts of the house showed how the larvae fit appropriate stones in the gaps, and cement them in place. Just as impressive is the log house at top right of the picture. This was built not by a caddis larva but by a caterpillar, a so-called bagworm. Caddises in water and bagworms on land have converged independently on the habit of building houses from materials that they gather from their surroundings. The picture shows a selection of caddis and bagworm houses.
If only chimps had the skills of a caddis larva…
The word ‘phenotype’ is used for the bodily manifestation of genes. The legs and antennae, eyes and intestines are all parts of the caddis’s phenotype. The gene’s-eye view of evolution regards the phenotypic expression of a gene as a tool by which the gene levers itself into the next generation – and, by implication, an indefinite number of future generations. What this chapter adds is the notion of the extended phenotype. Just as the shell of a snail is part of its phenotype, its shape, size, thickness, etc. being affected by snail genes, so the shape, size, etc. of a stone caddis house or twiggy bagworm cocoon are all manifestations of genes. Because these phenotypes are not part of the animal’s own body, I refer to them as extended phenotypes.
These elegant constructions must be the products of Darwinian evolution, no less than the armoured body wall of a lobster, a tortoise, or an armadillo. And no less than your nose or big toe. This means they have been put together by the natural selection of genes. Such is the Darwinian justification for speaking of extended phenotypes. There must be genes ‘for’ the various details of caddis and bagworm houses. This means only that there must be, or have been, genes in the insects’ cells, variants of which cause variation in the shape or nature of houses. To conclude this, we need assume only that these houses evolved by Darwinian natural selection, an assumption that no serious biologist would dispute, given their elegant fitness for purpose. The same is true of the nests of potter wasps, mud dauber wasps, and ovenbirds. Built of mud rather than living cells, they are extended phenotypes of genes in the bodies of the builders.
While their grasshopper cousins sing with serrated legs, male crickets sing with their wings, scraping the top of one front wing against a rough ‘file’ on the underside of the other front wing. Among their songs, the ‘calling song’ is loud enough to attract females within a certain radius, and to deter rival males. But what if it could be amplified, widening the catchment area for pulling females? Some kind of megaphone, perhaps? We use a megaphone as a simple directional amplifier, which works by ‘impedance matching’. No need to go into what that means, except to say that, unlike an electronic amplifier, it adds no extra energy. Instead, it concentrates the available energy in a particular direction. Could a cricket grow a megaphone out of its horny cuticle – a phenotype in the conventional sense? Like the remarkable backwards-facing trombone of the dinosaur Parasaurolophus, which probably served as a resonator for its bellowings. Crickets could have evolved something like that. But an easier material was to hand, and mole crickets exploited it.
CADDIS
BAGWORM
EXTENDED PHENOTYPES BUILT OF MUD
Potter wasp
Mud dauber
Ovenbird
Mole crickets, as their name suggests, are digging specialists. Their front legs are modified to form stout spades, strongly resembling those of moles, albeit on a smaller scale. The similarity, of course, is convergent. Some species of mole crickets are so deeply committed to underground life that they cannot fly at all. Given that a mole cricket could benefit from a megaphone, and given that it digs a burrow, what more natural than to shape the burrow as a megaphone? In the case of Gryllotalpa vineae it is a double megaphone, like an old-fashioned clockwork gramophone with two horns. Henry Bennet-Clark showed that the double horn concentrates the sound into a disc section rather than letting it dissipate in all directions as a hemisphere. Bennet-Clark was able to hear a single Gryllotalpa vineae (a species he discovered himself) from 600 metres away. The range of no ordinary cricket comes close.
Parasaurolophus
Assuming it’s as beautifully functional as it seems to be, the mole cricket’s megaphone must have evolved by natural selection, as a step-by-step improvement, in just the same way as the digging hand or as any part of the cricket’s own body. Therefore, there must be genes controlling horn shape, just as there are genes controlling wing shape or antenna shape. And just as there are genes controlling the patterning of cricket song itself. If there were no genes for horn shape, there would be nothing for natural selection to choose. Once again, remember that a gene ‘for’ anything is only ever a gene whose alternative alleles encode a difference between individuals.
Mole cricket
Mole
Mole cricket with double megaphone burrow
Now, when contemplating the double megaphone (or, for that matter, the houses of caddises and bagworms) you might be tempted to say something along the following lines. Cricket burrows are not like wings or antennae. They are the product of cricket behaviour, whereas wings and antennae are anatomical structures. We are accustomed to the idea of anatomical structures being under the control of genes. Can the same be said of behaviour, of cricket digging behaviour, or the sophisticated stonemasonry behaviour of a caddis larva? Yes, of course it can. And there is nothing to stop it being said of artifacts that are produced by the behaviour. The artifacts are just one further step in the causal chain from gene to protein to … a long cascade of processes in the embryo, culminating in the adult body.
There are numerous studies of the genetics of behaviour, including, as it happens, the genetics of cricket song. I want to discuss this work because, weirdly, behaviour genetics arouses a scepticism never suffered by anatomical genetics. Cricket song (though not specifically mole cricket song) has been the subject of penetrating genetic research by David Bentley, Ronald Hoy, and their colleagues in America. They studied two species of field cricket, Teleogryllus commodus from Australia and Teleogryllus oceanicus, also Australian but found in Pacific islands too. Adult crickets who have been brought up in isolation from other crickets sing normally. Nymphs who have not yet undergone their final moult to adulthood never sing, but in the laboratory their thoracic ganglia can be induced to emit nerve impulses with a time-pattern identical to the species song pattern. These facts strongly suggest that the instructions for how to sing the species song are coded in the genes. And those genes must be relevantly different in the two species, for their song patterns are different. This is beautifully confirmed by hybridisation experiments.
In nature these two Teleogryllus species don’t interbreed, but they can be induced to do so in the laboratory. The diagram, from Bentley and Hoy, shows the songs of the two species and of various hybrids between them. All cricket songs are made up of pulses separated by pauses. T.oceanicus (A in the picture) has a ‘chirp’ consisting of about five pulses followed by a series of about ten ‘trills’, each trill always made up of two pulses, closer to each other than the pulses of the chirp. We hear a rhythmic repetition pattern of trills. To my ears the trills sound slightly quieter than the chirps. After about ten of these double-pulse trills there’s another chirp. And the cycle repeats rhythmically, over and over again indefinitely. T.commodus (F) has a similar pattern of alternating chirps and trills. But instead of a series of ten or so double-pulse trills, there is only one long trill or perhaps two, between chirps.
Songs of pure bred and hybrid crickets
Now to the interesting question: what about the hybrids? Hybrid songs (C and D) are intermediate between those of the two parent species (A and F). It makes a difference which species is the male (compare C with D), but we needn’t go into that here, interesting though it is for what it might tell us about sex chromosomes. In any case, hybrid song is a beautiful confirmation of genetic control of a behaviour pattern. Further evidence (B and E) comes from crossing hybrids with each of the two wild species (what geneticists call a backcross). If you compare all five songs, you’ll note a satisfying generalisation: hybrid songs resemble the two wild species’ songs in proportion to the number of genes the hybrid individual has inherited from each species. The more oceanicus genes an individual has, the more its song resembles wild oceanicus rather than commodus. And vice versa. As your eyes move down the page from oceanicus towards commodus, the more you detect resemblance to commodus song. This suggests that several genes of small effect (‘polygenes’) sum their effects. And what is not in doubt is that the species-specific song patterns that distinguish these two species of crickets are coded in the genes: a nice example of how behaviour is just as subject to genetic control as anatomical structures are. Why on earth shouldn’t it be? The logic of gene causation is identical for both. Both are products of a chain of causation, with the behaviour having one more link in the chain.
You could do a similar study of the genetics of megaphone-building behaviour. But you might as well go to the next step in the causal chain, the megaphone itself. Do a genetic study of differences between megaphones. They are extended phenotypes of mole cricket genes. This has not been done, but nothing prevents it. Again, nobody has studied the genetics of caddis houses, but there’s no reason why they shouldn’t, although there might be practical difficulties in breeding them in the lab. Michael Hansell was once giving a talk at Oxford, on the building behaviour of caddis larvae. In passing, he was lamenting his failed attempts to breed caddises in the lab, for he wished he could study their genetics. At this, the Professor of Entomology growled from the front row: ‘Haven’t you trrrried cutting their heads off?’ It seems that the insect brain exercises inhibitory influences such that beheading can be expected to have a releasing effect.
If you were to succeed in breeding caddises in captivity, you could systematically select changes in caddis houses over generations. Or you could artificially select for mole cricket megaphone size or shape, generation by generation, breeding from those individuals whose horns happen to be wider, or deeper, or of a different shape. You could breed giant megaphones, just as you might breed giant antennae or mandibles.
That would be artificial selection, but something like it must have happened through natural selection. Whether by artificial or natural selection, the evolution of larger megaphones could come about only by differential survival of genes for megaphone size. For the megaphone to have evolved in the first place as a Darwinian adaptation, there had to be genes for megaphone shape. The notion of the extended phenotype is a necessary part of the gene’s-eye view of evolution. The extended phenotype should be an uncontroversial addition to Darwinian theory.
But aren’t those ‘genes for megaphone shape’ really genes for altered digging behaviour, which is part of the ‘ordinary’ phenotype of the cricket? Aren’t genes for caddis house shape ‘really’ genes for building behaviour, that is to say, ‘ordinary’ phenotypic manifestations within the body? Why talk about ‘extended’ phenotypes outside the body at all? Well, you could equally well say that the genes for altered digging behaviour are ‘really’ genes for changed wiring in the ganglia in the thorax. And genes for changes in the thoracic ganglia are, in turn, ‘really’ genes for changes in cell-to-cell interactions in embryonic development. And they, in turn, are ‘really’ … and so on back until we hit the ultimate ‘really’. Genes are really really really only genes for changed proteins, assembled according to the rules for translating the sixty-four possible DNA triplet codons into twenty amino acids plus a punctuation mark. I repeat, because it is important, we have here a chain of causation whose first steps (DNA codons choosing amino acids) are knowable, whose final step (megaphone shape) is observable and measurable, and whose intermediate steps are buried in the details of embryology and nerve connections – perhaps inscrutable but necessarily there. The point is that any one of those many intermediate steps in the chain of causation could be regarded as ‘phenotype’, and could be the target of selection, artificial or natural. There is no logical reason to stop the chain at the animal’s body wall. Megaphone is ‘phenotype’, every bit as much as nerve-wiring is phenotype. Every one of those steps, both in the cricket’s body and extended outside it, can be regarded as caused by gene differences. Just the same is true of the chain of causation leading from genes to caddis house, even though the behavioural step, the actual building itself, involves sophisticated trial and error in the selection of suitable stones and rotating them into position to fit the existing structure. And now to advance the argument a stage further. The extended phenotype of a gene can reach into the body of a different individual.
Natural selection doesn’t see genes for digging behaviour directly, nor does it see neuron circuitry directly, nor indeed megaphone shape directly. It sees, or rather hears, song loudness. Gene selection is what ultimately matters, but song loudness is the proxy by which gene selection is mediated, via a long series of intermediates. But even song loudness is not the end of the causal chain. As far as natural selection is concerned, song loudness only matters insofar as it attracts females (and deters males, but let’s not complicate the argument). The causal chain extends to a radius where it exerts an influence on a female cricket. This has to mean that a change in female behaviour is part of the extended phenotype of genes in a male cricket. Therefore, the extended phenotype of a gene can reside in another individual. The general point I am aiming towards is that the phenotypic expression of a gene can extend even to living bodies other than the body in which the genes sit. Just as we can talk of a gene ‘for’ a Hapsburg lip, or a gene ‘for’ blue eyes, so it is entirely proper to talk of a gene (in a male cricket) ‘for’ a change in another individual’s behaviour (in this case a female cricket).
We saw in Chapter 7 that song in male canaries and ring doves has a dramatic effect on female ovaries. They swell hugely, with a corresponding rush of hormones and all that it entails. The consequent changes in female behaviour and physiology are in truth phenotypic expression of male genes. Extended phenotypic expression. You may deny it only if you deny Darwinian selection itself.
Ears are not the only portals into a female dove’s brain through which a male’s genes might exert an extended phenotypic influence. Male birds of many species glow with conspicuous colours. These cannot be good for individual survival, but they are still good for the survival of the genes that fashioned them. They achieve this good by assisting individual reproduction at the expense of individual survival. With few exceptions, it is males that sacrifice their personal longevity on the altar of gene survival, through sexually attractive coloration. In those species such as pheasants or birds of paradise, where males dazzle, females are usually drabber in colour, often well camouflaged. Bright coloration in males is favoured, either through attracting females or through besting rival males. In both cases, the naturally selected genes for bright coloration have extended phenotypic expression in the changed behaviour of other individuals. I don’t know whether exposure to a male peacock fan causes peahen ovaries to change, as male dove bow-cooing song does to female dove ovaries. It wouldn’t surprise me. I’d even be surprised if it didn’t.
Unfortunately, predators tend to have eyes like the eyes of the females whom the male is seeking to impress. What is conspicuous to one will probably be conspicuous to all. It’s worth it to the male, or rather to the genes that coloured him. Even if his finery costs him his life, it can already have paid its way in previous success with females. But is there some way a male bird could manipulate females via their eyes without calling attention to himself? Could he shed his dangerously conspicuous personal phenotype, offloading it to an extended phenotype at a safe distance from his own body? ‘Shed’ and ‘offload’, of course, must be understood over evolutionary time. We aren’t talking about shedding feathers in an annual moult, although that happens too – perhaps for the same reason. Black-headed gulls, for instance, shed their conspicuously contrasting face masks as soon as the breeding season is over.
Bower birds are a family of birds inhabiting the forests of New Guinea and Australia. Their name comes from a remarkable and unique habit. They build ‘bowers’ to seduce females. The skills needed to build a bower could be seen as a distant derivative of nest-building skills, and perhaps ultimately derived from them. But the bower is emphatically not a nest. No eggs are laid in it, no chicks reared there. Female bower birds build nests to house eggs as other birds do, and their nests don’t resemble male bowers.
The bower’s sole purpose is to attract females, and males take enormous pains in their creation. First, they clear stray leaves and other debris from the arena in which the bower is to be built. Then the bower itself is assembled from twigs and grass. The details vary from species to species. Some resemble a Robinson Crusoe hat, some a grand archway, others a tower. The final stage of bower design is, I think, the most remarkable of all. The ground in front of and under the bower is colourfully and – I can’t resist saying – tastefully decorated. The male gathers decorative objects – coloured berries, flowers, even bottle tops. Movies of male bower birds at work irresistibly remind me of an artist putting the finishing touches to a canvas, standing back, head cocked judgmentally, then darting forward to make a delicate adjustment, standing back again and surveying the effect with head on one side before darting forward again. That is what emboldened me to use a word like ‘tastefully’. It is hard to resist the impression that the bird is exercising his aesthetic judgement in perfecting a work of art. Even if the decorated bower is not to every human’s taste, or even every female bower bird’s, the ‘touching up’ behaviour of the male almost forces the conclusion that the male has taste of his own, and he is adjusting his bower to meet it.
Remember the discussion in Chapter 7, where I suggested that when male songbirds learn to sing, they are exercising their own aesthetic judgement? The evidence shows, you’ll remember, that young birds burble at random, choosing, by reference to a template, which random fragments to incorporate into their mature song. The male, I argued, has a similar brain to a female of his own species. Not surprisingly, therefore, whatever appeals to him can be expected to appeal to her. The development of song in the young bird could be regarded as a work of creative composition in which the male adopts the principle of ‘whatever turns me on will probably appeal to a female too’. I see no reason to refrain from a similar aesthetic interpretation of bower-building. ‘I like the look of a heap of blue berries just there. So there’s a good chance that a female of my own species will like it too … And perhaps a single red flower over there … or, no, it looks better here … and better still, slightly to the left, and why not set it off with some red berries?’ Of course, I am not literally suggesting that he thinks it through in so many words.
Species differ as to their preferred decoration colours, as well as the shape of their bowers. The satin bower bird (here) goes for blue, a fact that may be connected with the blue-black sheen of his plumage, or the species’ brilliant blue eyes. The male satin bower bird who built this bower has discovered blue drinking straws and bottle tops, and laid out a rich feast of blue to delight the female eye. More soberly, the Great Bower Bird, Chlamydera nuchalis, says it with shells and pebbles (opposite).
The bower is an extended phenotype of genes in the body of the male bower bird. An external phenotype, which presumably has the advantage that its extravagance is not worn on the body and therefore will not call predators’ attention to the male himself. I do not know whether exposure to a more than usually magnificent bower stimulates a hormone surge in the blood of a female, but again the research on ring doves and canaries would lead me to expect this.
We are accustomed to thinking of genes as being physically close to their phenotypic targets. Extended phenotypes can be large, and far distant from the genes that cause them. The lake flooded by a beaver’s dam is an extended phenotype of beaver genes, extended in some cases over acres. The songs of gibbons can be heard a kilometre away in the forest, howler monkeys as much as five kilometres: true genetic ‘action at a distance’. These vocalisations have been favoured by natural selection because of their extended phenotypic effect on other individuals. Chemical signals can achieve a great range among moths. Visual signals require an uninterrupted line of sight, but the principle of genetic action at a distance remains. The gene’s-eye view of evolution necessarily incorporates the idea of the extended phenotype. Natural selection favours genes for their phenotypic effects, whether or not those phenotypic effects are confined to the body of the individual whose cells contain the genes.
In 2002, Kim Sterelny, editor of the journal Biology and Philosophy, marked the twentieth anniversary of the publication of The Extended Phenotype by commissioning three critical appraisals, plus a reply from me. The special issue of the journal came out in 2004. The criticisms were thoughtful and interesting, and I tried to follow suit in my reply, but all this would take us too far afield here. I concluded my piece with a humorously grandiose fantasy about the building of a future Extended Phenotypics Institute. This pipedream edifice was to have three wings, the Zoological Artifacts Museum (ZAM), the laboratory of Parasite Extended Genetics (PEG), and the Centre for Action at a Distance (CAD). The subjects covered by ZAM and CAD have dominated this chapter. PEG must wait till the final chapter. Parasites often exert dramatic extended phenotypic effects on their hosts, manipulating the host’s behaviour to the parasite’s advantage, often in bizarrely macabre ways. The parasite doesn’t have to reside in the body of the host, so there is an overlap with CAD, the Action at a Distance wing. Cuckoo chicks are external parasites who exert extended phenotypic influence over the behaviour of their foster parents. And cuckoos are so fascinating they deserve a chapter of their own. For a different reason, now to be explained.
10 The Backward Gene’s-Eye View
The previous two chapters constituted my short reprise of the gene’s-eye view of evolution as I explained it in The Selfish Gene and The Extended Phenotype. I want, now and in the next chapter, to offer the gene’s-eye view in another way, a way that is particularly suitable for The Genetic Book of the Dead. This is to imagine the view seen by a gene as it ‘looks’ backwards at its ancestral history. A vivid example concerns the cuckoo. To which deplorable bird we now turn.
‘Deplorable bird’? Of course I don’t really mean that. The phrase amused me in a Victorian bird book belonging to my Cornish grandparents, where it referred to the cormorant. Each page of the book was devoted to one species. When you turned to the cormorant’s page, the very first sentence to greet you was, ‘There is nothing to be said for this deplorable bird.’ I can’t remember what grudge the author held against the cormorant. He might have had better grounds with the cuckoo, which is certainly deplorable from the point of view of its foster parents but, as a Darwinian biologist, I think it is a supreme wonder of the world. ‘Wonder’, yes, but there’s also an element of the macabre in the spectacle of a tiny wren devotedly feeding a chick big enough to swallow it whole.
Everyone knows that cuckoos are brood parasites who trick nesting birds of other species into rearing their young. ‘Cuckoo in the nest’ is proverbial. John Wyndham’s The Midwich Cuckoos, about aliens implanting their young in unwitting human wombs, is one of several works of fiction whose titles sound the cuckoo motif. Then there are cuckoo bees, cuckoo wasps, and cuckoo ants who, in their own hexapod ways, hijack the nurturing instincts of other species of insect. The cuckoo fish, a kind of catfish from Lake Tanganyika, drops its eggs among the eggs of other fish. In this case the hosts are ‘mouthbreeders’, fish belonging to the Cichlid family who take their eggs and young into their own mouths for protection. The cuckoo fish’s eggs and later fry are welcomed into the unsuspecting host’s mouth, and tended as lovingly as the mouthbreeder’s own.
Plenty of bird species have independently evolved their own versions of the cuckoo habit, for example the cowbirds of the New World, and cuckoo finches of Africa. Within the cuckoo family itself (Cuculidae), 59 of the 141 species parasitise other species’ nests, the habit having evolved there three times independently. In this chapter, unless otherwise stated, for the sake of brevity I use the name cuckoo to mean Cuculus canorus, the so-called common cuckoo. Alas, it’s not common anymore, at least in England. I miss their springtime song even if their victims don’t, and was delighted to hear it on a recent visit to a beautiful, remote corner of western Scotland where it ‘shouts all day at nothing’. My main authority – indeed today’s world authority – is Professor Nick Davies of Cambridge University. His book Cuckoo is a delightful amalgam of natural history and memoir of his field research on Wicken Fen, near Cambridge. Described by David Attenborough as one of the country’s greatest field naturalists, he achieves heights of lyrical word-painting unsurpassed in the literature of modern natural history:
North towards the horizon is the eleventh-century cathedral of Ely, which sits on the raised land of the Isle of Ely, from where Hereward led his raids against the Normans. In the early mornings, when the mist lies low, the cathedral appears as a great ship, sailing across the fens.
The ruthlessness of the cuckoo begins straight out of the egg. The newly hatched chick has a hollow in the small of the back. Nothing sinister about that, you might think. Until you are told the sole use to which it is put. The cuckoo nestling needs the undivided attention of its foster parents. Rivals for precious food must be disposed of without delay. If it finds itself sharing the nest with either eggs or chicks of the foster species, the hatchling cuckoo fits them neatly into the hollow in its back. It then wriggles backwards up the side of the nest and tosses the competing egg or chick out. There is, of course, no suggestion that it knows what it’s doing, or why it is doing it, no feelings of guilt or remorse (or triumph) in the act. The behavioural routine simply runs like clockwork. Natural selection in ancestral generations favoured genes that shaped nervous systems in such a way as to play out this instinctive act of (foster) fratricide. That is all we can say.
And there’s no more reason to expect the foster parents to know what they are doing when they fall for the cuckoo’s trick. Birds are not little feathered humans, seeing the world through the lens of intelligent cognition. It makes at least as much sense to see the bird as an unconscious automaton. This helps us understand the otherwise surprising behaviour of foster parents. A pioneering cinematographer of the cuckoo’s dark ways was Edgar Chance, avid ornithologist of the early twentieth century. By Nick Davies’s account of his film, a mother meadow pipit appeared totally unconcerned as it watched its own precious offspring being murdered by the cuckoo chick in its nest. The mother then left on a foraging trip, as if nothing untoward had happened. When she returned, she pointlessly fed her chick as it lay dying on the ground. From a human cognitive point of view, her behaviour makes no sense: neither the impassive watching of the initial murder nor the subsequent futile feeding of the doomed chick. We shall meet this point again and again throughout the chapter.
The name ‘cuckoo’ is derived from the simple, two-note tune of the male bird’s song, so simple indeed that some ornithologists downgrade it from ‘song’ to ‘call’ (on parallel grounds to the hysterically unpopular downgrading of Pluto to sub-planet status). The cuckoo’s song (or call) is commonly described as dropping through a minor third, but I’m happy to quote no less an authority than Beethoven in support of my hearing it as a major third. His famous cuckoo in the Pastoral Symphony descends from D to B Flat. Whether major or minor, whether song or call, it is simple – and perhaps has to be simple because the male never gets a chance to learn it by imitation. A cuckoo never meets either biological parent. It knows only its foster parents, who could belong to any of a variety of species, each with its own song, which the young cuckoo must not learn. So the male cuckoo’s song has to be hard-wired genetically, and a kind of common sense concludes, not very confidently, that it should therefore be simple.
Now we approach the remarkable story that earns the cuckoo its place in a chapter on genes ‘looking backwards in time’. Cuckoo eggs mimic the colour and patterning of the other eggs in the particular foster nest in which they sit. And they mimic them even though many different foster species are involved, with very different eggs. Here is a clutch of six brambling eggs plus one cuckoo egg. The only way I, and doubtless you, can tell which one is the cuckoo egg is by its slightly larger size.
At first sight, such egg mimicry might seem no more remarkable than the ‘paintings’ of Chapter 2. Well, that’s quite remarkable enough! But now look at the next picture showing a parasitised nest of meadow pipit eggs.
Again, you can spot the tell-tale size of the cuckoo egg. But what is really noticeable is that the cuckoo egg in the second picture is dark with black speckles like meadow pipit eggs, whereas the cuckoo egg in the first picture is light and with rusty speckles like brambling eggs. Meadow pipit eggs are dramatically different from brambling eggs. Yet cuckoo eggs achieve a near-perfect colour match in each of the two nests.
Once again, the mimicry might seem par for the course, all of a piece with the lizard, frog, spider, or ptarmigan ‘paintings’ of Chapter 2. It would indeed be relatively unremarkable if the cuckoos that parasitise bramblings were a different species from the cuckoos that parasitise meadow pipits. But they aren’t. They’re the same species. Males breed indiscriminately with females reared by any foster species, so the genes of the whole species are mixed up as the generations pass. That mixing is what defines them all as of the same species. Different females, all belonging to the same species and consorting with the same males, parasitise redstarts, robins, dunnocks, wrens, reed warblers, great reed warblers, pied wagtails, and others. But each female parasitises only one of those host species. And the remarkable fact is that (with a few revealing exceptions) the eggs of each female cuckoo faithfully mimic those of the particular host in whose nest she lays them. The only consistent betrayer is that cuckoo eggs are slightly larger than the host eggs that they mimic. Even so, they are smaller than they ‘should’ be for the size of the cuckoo itself. Presumably, if the pressure to mimic drove them to be any smaller, the chicks would be penalised in some way. The actual size is a compromise between pressure to be small to mimic the host eggs, and an opposite pressure towards the larger optimum for the cuckoo’s own size.
I doubt that you’re wondering why egg mimicry benefits the cuckoos. Foster parents are mostly very good at spotting cuckoo eggs, and they often eject them. A cuckoo egg of the wrong colour would stand out like a sore thumb. Actually, that’s an unusually poor cliché. Have you ever seen a sore thumb, and did it stand out? Let’s initiate a new simile. Stands out like a baseball at Lord’s? Like a Golden Delicious in a basket of genuinely delicious apples? Just look at that cuckoo egg in the brambling nest and imagine transplanting it into the meadow pipit nest. Or vice versa. The host birds would unhesitatingly toss it out. Or, if tossing it out is too difficult, abandon the nest altogether. Such discrimination is not a surprise when you consider that bird eyes are acute enough to perfect the exquisitely detailed painting of lichen-mimicking moths and stick-mimicking caterpillars.
Foster parents, then, whether as automata or cognitively, can be expected to provide the selection pressure that explains why it might benefit cuckoo eggs to show such beautiful egg mimicry. They throw out eggs that don’t look like their own. But what is surprising, hugely so, is that cuckoos, all of one intrabreeding species, manage to mimic the eggs of many different foster parent species. To drive home the point, here’s yet another example: a reed warbler nest with, once again, wonderful egg mimicry by the single, slightly larger cuckoo egg.
These beautiful examples force us back to the central question of this whole discussion. How is it possible for female cuckoos, all belonging to the same species and all fathered by indiscriminate males, to produce eggs that match such a range of very different host eggs? Are we to believe that female cuckoos take one look at the eggs in a nest and take a decision to switch on some kind of alternative egg-colouring mechanism in the lining of the oviduct? That is improbable, to say the least. There are women who might love to control, by sheer willpower and for very different reasons, the behaviour of their own oviduct. But it’s not the kind of thing willpower does. And, with the best will in the world, it’s not clear how will will power it.
What is the true explanation for the female cuckoo’s apparent versatility? Nobody knows for sure, but the best available guess makes use of a peculiarity of bird genetics. As you know, we mammals determine our sex by the XX / XY chromosome system. Every woman has two X-chromosomes in all her body cells, so all her eggs have an X-chromosome. Every man has an X- and a Y-chromosome in all his body cells. Therefore, half his sperms are Y sperms (and would father a son when coupled with a necessarily X egg) and half are X sperms (would father a daughter when coupled with a necessarily X egg). Less well known is that birds have a similar system, but it evidently arose independently because it is reversed. The chromosomes are called Z and W instead of X and Y, but that’s not important. What matters is that in birds females are ZW and males are ZZ. That’s opposite to the mammal convention, but otherwise the principle is the same. Whereas the Y-chromosome passes only down the male line in mammals, in birds the W chromosome passes only down the female line. The W comes from the mother, the maternal grandmother, the maternal maternal great grandmother and so on back through an indefinite number of female generations.
Now recall the title of this chapter: ‘The Backward Gene’s-Eye View’. It’s all about genes looking back at their own history. Imagine you are a gene on the W-chromosome of a cuckoo, looking back at your ancestry. Not only are you in a female bird today, you have never been in a male bird. Unlike the other genes on ordinary chromosomes (autosomes), which have found themselves in male and female bodies equally often down the ages, the ancestral environments of the W-chromosome have been entirely confined to female bodies. If genes could remember the bodies they have sat in, the memories of W-chromosomes would be exclusively of female bodies not male ones. Z-chromosomes would have memories of both male and female bodies.
Hold that thought while we look at a more familiar kind of memory: memory by the brain, individual experience. It is a fact that female cuckoos remember the kind of nest in which they were reared, and choose to lay their own eggs in nests of the same foster species. Unlike the improbable feat of controlling your own oviduct, remembering early experience is exactly the kind of thing bird brains are known to do. When they come to choose a mate, as we saw in Chapter 7, birds of many species refer back to a kind of mental photograph of their parent, which they filed away in memory after their first encounter on hatching (‘imprinting’): even if – in the case of incubator-hatched goslings, for instance – what they later find attractive is Konrad Lorenz. To remember Lorenz, parental plumage, father’s song, or foster-parent’s nest – it’s all the same kind of problem. The same imprinting brain mechanism works well enough in nature even if, in captivity, it misfires.
I think you can see where this argument is going. Each female mentally imprints on the same foster nest as her mother; and therefore her maternal grandmother; and her maternal maternal great grandmother. And so on back. And her childhood imprinting leads her to choose the same kind of nest as her female forebears. So, she belongs to a cultural tradition going exclusively down the female line. Among females there are robin cuckoos, reed warbler cuckoos, dunnock cuckoos, meadow pipit cuckoos, etc., each with their own female tradition. But only females belong to these cultural traditions. Each cultural line of females is called a gens – plural gentes. A female may belong to the meadow pipit gens, or the robin gens, or the reed warbler gens, etc. Males don’t belong to any gens. They are descended from – and they father – females of all gentes indiscriminately.
Finally, we put these two strands of thought together, again in the light of the chapter’s title. With the exception of W-chromosome genes, all the genes in a female cuckoo look back through a chain of ancestors belonging to every gens that’s going. W-chromosomes aside, gentes are not genetically separate like true races, because males confound them. Only W-chromosome genes are gens-specific. Only W-chromosomes look back on ancestors of a particular gens to the exclusion of any other. We talked of two kinds of memory: genetic memory and brain memory. See how the two coincide where W-chromosome genes are concerned!
With respect to the W-chromosome, and only the W-chromosome, gentes are separate genetic races. So – I think you’ve already completed the argument yourself – if the genes that determine egg coloration and speckling are carried on the W-chromosome, it would solve the riddle we began with, the riddle of how it’s possible for the females of one species of cuckoo to mimic the eggs of a wide variety of host species. It isn’t willpower that chooses egg colour, it’s W-chromosomes.
You will have guessed that it’s not as simple as that. Things seldom are in biology. Although female cuckoos have a strong preference for their natal nest type when they come to lay, they occasionally make a mistake and lay in the ‘wrong’ nest, different from their natal nest. Presumably that’s how new gentes get their start. And not all gentes achieve good egg mimicry. Dunnock (hedge sparrow) eggs are a beautiful blue. But cuckoo eggs in dunnock nests aren’t blue (left). They aren’t even ‘trying’ to be blue, we might say. The cuckoo egg in the picture stands out like a sore … like a bloodhound in a pack of dachshunds. Are cuckoos, perhaps, constitutionally incapable of making blue eggs? No. Cuculus canorus in Finland has achieved a most beautiful blue, in perfect mimicry of redstart eggs (right). So why don’t cuckoo eggs mimic dunnock eggs? And how do they get away with it? The answer is simple, although it remains puzzling. Dunnocks are among several species that don’t discriminate, don’t throw out cuckoo eggs. They seem blind to what looks to us glaringly obvious. How is this possible, given that other small songbirds have powers of discrimination acute enough to perfect the finishing touches to the egg mimicry achieved by their respective gentes of female cuckoos? And given that bird eyes are capable of perfecting the detailed mimicry of stick caterpillars, lichen-mimicking moths, and the like?
Cuckoos and their hosts, as with stick caterpillars and their predators, are engaged in an ‘evolutionary arms race’ with one another. As mentioned in Chapter 4, arms races are run in evolutionary time. It’s a persuasive parallel to human arms races, which are run in ‘technological time’, and a lot faster. The aerial swerving and dodging chases of Spitfires and Messerschmitts were run in real time measured in split seconds. But in the background and more slowly, in factories and drawing-offices in Britain and Germany, races were run to improve their engines, propellers, wings, tails, weaponry, etc., often in response to improvements on the other side. Such technological arms races are run over a timescale measured in months or years. The arms races between cuckoos and their various host species have been running for thousands of years, again with improvements on each side calling forth retaliatory improvements in the other.
Nick Davies and his colleague Michael Brooke suggest that some gentes have been running their respective arms races for longer than others. Those against meadow pipits and reed warblers are ancient arms races, which is why both sides have become so good at outdoing the other – and therefore why the cuckoo eggs are such good mimics. The arms race against dunnocks, they suggest, has only just begun. Not enough time for the dunnocks to evolve discrimination and rejection. And not enough time for the dunnock gens of cuckoos to evolve the appropriate blue colour.
If it’s true that cuckoos have only just ‘moved into’ dunnock nests, we must suppose that these ‘pioneer’ cuckoos have ‘migrated’ from another host species, presumably one with rusty-spotted grey eggs because that’s the egg colour of the ‘newly arrived’ dunnock gens of cuckoo. I suppose this is how any new gens gets its start. But don’t be misled by ‘pioneer’ and ‘migrated’. It would not have been any kind of bold decision to sally forth into fresh nests and pastures new. It would have been a mistake. As we’ve seen, cuckoos do indeed occasionally lay an egg in the wrong kind of nest, a nest appropriate to a different gens. Their egg then really does stand out like a … invent your own substitute for the sore thumb cliché. Natural selection normally penalises such blunders, we can presume, pretty promptly. But what if it’s a new host species that hasn’t yet been ‘invaded’ by cuckoos. The new host species is naive. They haven’t hitherto had any reason to throw out mismatched eggs. Once again, remember, birds are not little feathered humans with human judgement. The arms race has yet to get properly under way. And the host species can expect to remain naive while the arms race is yet young. But how young is young? Strangely enough, we are not totally without evidence bearing on the question, as Nick Davies points out.
Call the witness Geoffrey Chaucer. In The Parlement of Foules (1382), the cuckoo is reproached: ‘Thou mordrer of the heysugge on the braunche that broghte thee forth.’ Another name for dunnock is hedge sparrow or, in Middle English, heysugge (heysoge, heysoke, eysoge). This would seem to suggest that cuckoos were already parasitising dunnocks in the fourteenth century, when Chaucer wrote. Is 650 years long enough for an arms race to reach some sort of perfection of mimicry? Perhaps not, given that, as Davies points out, only 2 per cent of dunnock nests are parasitised. Maybe, then, the selection pressure is so weak that a 600-year-old arms race is indeed young.
I prefer to add two further suggestions. The first concerns identification. Did Chaucer really mean dunnock when he said heysugge? When we say ‘sparrow’ we normally mean the house sparrow, Passer domesticus, not the hedge sparrow or dunnock, Prunella modularis. Yet the English word ‘sparrow’ is used for both. To many who are not avid twitchers, all little brown birds (LBBs) look much the same, and we might even sink so low as to call them all ‘sparrows’. I can’t help wondering whether Chaucer was using ‘heysugge’ to mean LBB rather than specifically Prunella modularis?
My second suggestion is more biologically interesting. If we think carefully about it, there’s no reason, is there, to suppose that there’s only one cuckoo gens for each host species? Maybe Chaucer’s gens of dunnock cuckoos has died out, and a new gens of dunnock cuckoos is just beginning its arms race. Perhaps other gentes of dunnock cuckoos have perfect egg mimicry today, but have not come to the notice of ornithologists. There would be no relevant gene flow between them because males don’t have W-chromosomes.
Claire Spottiswoode and her colleagues are running a parallel study of an unrelated South African finch, which convergently evolved the cuckoo habit. The cuckoo finch, Anomalospiza imberbis, lays its eggs in the nests of grass warblers. Different gentes of cuckoo finch mimic the eggs of different grass warbler species. There is genetic evidence that what distinguishes the gentes is indeed their W-chromosomes, which reinforces the idea that the same thing is going on in cuckoos. As Dr Spottiswoode points out, this doesn’t have to mean that every detail of all the egg colours is carried on the W-chromosome. In both cuckoos and cuckoo finches, genes for making all the different egg colours have very probably been built up on other chromosomes (‘autosomes’) over many generations, and are carried by all the gentes and passed on by males as well as females. The W-chromosome need only have switch-genes – genes that switch on or off whole suites of genes carried on autosomes. And the relevant autosomal genes would be carried by males as well as females.
This is indeed how sex itself is determined. If you have a Y-chromosome, you have a penis. If you have no Y-chromosome, you have a clitoris instead. But there’s no reason to suppose that the genes that influence the shape and size of a penis are confined to the Y-chromosome. Far from it. It’s entirely plausible that they are scattered over many autosomes. There’s no reason to doubt that a man may inherit genes for penis size from his mother as well as from his father. Presence or absence of a Y-chromosome determines only which alternative suite of genes on autosomes will be switched on. For most purposes you can think of the entire Y-chromosome as a single gene that switches on suites of other genes on autosomes elsewhere in the genome. A point of terminology: members of these suites of autosomal genes are called ‘sex-limited’ as distinct from ‘sex-linked’. Sex-linked genes are those that are actually carried on the sex-chromosomes themselves.
Probably the best guess towards a solution of the riddle of cuckoo egg mimicry is that suites of genes on lots of chromosomes determine egg coloration and spotting. These are equivalent to ‘sex-limited’, and we may call them ‘gens-limited’. They are switched on or off by the presence or absence of one or more genes on the W-chromosome, genes that, by analogy, we can call ‘gens-linked’. All cuckoo autosomes may have suites of genes for mimicking a whole repertoire of host eggs. W-chromosomes contain switch genes that determine which suite of genes is turned on. And it is W-chromosomes that are peculiar to each gens of females, W-chromosomes that look back at their history and see a long line of nests of only one foster species.
This interpretation of egg mimicry in cuckoos is my introduction to the topic of the backward gene’s-eye view, genes looking over their shoulder at their own ancestry. Here’s a similar but more complicated example involving fish and the Y-chromosome. Different kinds of fish display a bewildering variety of sex-determining systems. Some don’t use sex chromosomes at all but determine sex by external cues. Some fish are like birds in that females are XY and males are XX. Others are like us mammals: males are XY, females XX. Among these are small fish of the genus Poecilia, which includes mollies and guppies among popular aquarium fish. One species, Poecilia parae, has a remarkable colour polymorphism, which affects only the males. Polymorphism means that there are different genetically determined colour types coexisting in the population (in this case five colour patterns) and the proportions of the different types remain stable in the population through time. All five male morphs can be found swimming together in South American streams. There’s only one female morph: females look alike.
Since the polymorphism affects only one sex, we can call them five gentes, by analogy with the cuckoos, with the difference that in these fish it’s the males who are separated by gens. The picture shows the five male types plus a female at the bottom. Three of the five male types have two long stripes like tramlines. Between the tramlines there is colour, and I’ll call them reds, yellows, and blues respectively. These three ‘tramliners’ can, for many purposes, be lumped together. The fourth type has vertical stripes. They’re officially named ‘parae’, but confusingly that’s also the name of the whole species. I’ll call them ‘tigers’. The fifth type, ‘immaculata’, is relatively plain grey, like females but smaller, and I’ll call them ‘greys’.
Tigers are the largest. They behave aggressively, chasing rival males away, and copulating with females by force. Greys are the smallest, and they manage to copulate only by occasionally sneaking up on females opportunistically. When they get away with it, it seems to be because otherwise aggressive males mistake them for females, which they do indeed resemble. Greys have the largest testes, presumably capable of producing the most sperm, perhaps to take advantage of their scarce opportunities to use it. Red, yellow, and blue tram-liners are of intermediate size. Rather than rape or sneak, they court females in a civilised manner, displaying their respective coloured flanks.
Tiger
Grey
Blue
Yellow
Red
Female
Male ‘gentes’ in fish?
Now here’s where the parallel to cuckoos kicks in. Evidence suggests that colour morph inheritance runs entirely down the male line. In every case studied, sons belong to the same type as their father, and therefore paternal grandfather, paternal paternal great grandfather, etc. Their mother has no genetic say in the matter, and nor does their maternal grandfather, etc., even though each one belongs to one colour gens or another. This suggests the hypothesis that the five types of males differ with respect to their Y-chromosomes – just as gens-inheritance in female cuckoos seems to be carried on the W-chromosome. The details of colour pattern and behaviour of the male fish may be carried in suites of genes on autosomes (gens-limited). But the genes determining which gens an individual belongs to (and presumably which suite of colour and pattern genes on other chromosomes is switched on) seem to be gens-linked, that is, carried on the Y-chromosome.
Researchers are doing fascinating work on mate choice in these fish and are homing in on what maintains the polymorphism. It seems that each of the five male types has an equilibrium frequency, fitting the definition of a true polymorphism. If its frequency falls below the equilibrium, it is favoured and therefore becomes more frequent in the population. If its frequency rises too high, it is penalised and its frequency decreases. This so-called ‘frequency-dependent selection’ is a known way for polymorphisms to be maintained in a population. How might it work in practice? The details are not yet clear but might look something like this. The grey sneakers benefit from being mistaken for females. If they become too frequent, perhaps the real females or aggressive tigers get ‘wise’ to them. How about the tigers themselves? If they get too frequent, they waste time fighting each other instead of mating. This might give the greys more opportunity to sneak matings. As for the three ‘tramliners’, who court females in a gentlemanly manner by flashing their vividly coloured flanks, there is some evidence that females prefer rarer types. This would fit the ‘equilibrium frequency’ idea, although it’s not clear why females should exhibit such a preference. More research is needed and is under way now. I am grateful to Dr Ben Sandkam, formerly of the University of British Columbia and now at Cornell, for sharing with me his thoughts on these matters.
Now let’s again apply the backward-looking technique of this chapter. Every male of Poecilia parae can look back through a long line of male ancestors, all belonging to the same gens as him, and all sharing the same Y-chromosome. This is what makes it possible for suites of genes for colour patterning and associated behaviour to become switched on in separate gentes of males, despite their sharing the same ancestors in the female line. The gene’s-eye view of the past comes into its own again, as with the cuckoos. Autosomal genes, governing characteristics other than gens-specific colour, look back on ancestors of all gentes.
Returning to cuckoos, the ‘looking back’ ploy can help us answer another riddle, and it’s an even tougher one. Although most host species are very good at distinguishing cuckoo eggs from their own (how else could natural selection have perfected cuckoo egg mimicry?), they turn out to be lamentably bad later, failing to notice that the growing cuckoo fledgling is an impostor. Even though it dwarfs them, in most cases grotesquely so. A tiny warbler is in danger, you might think, of being swallowed whole by its monstrous foster child. Foster parents, of whatever species, end up dwarfed by the cuckoo nestling into whom they tirelessly shovel food, working every devoted daylight hour to do so. How do the cuckoo nestlings get away with such a transparent, over-the-top deception? Once again, we have to be more than usually on our guard against anthropomorphism. Do not ask whether the bird’s behaviour makes sense from a human-like cognitive perspective. Of course it doesn’t. Ask instead about selection pressures acting on ancestral genes that control the development of behavioural automatisms.
A warbler feeding a cuckoo
Even given this preliminary, I must admit that available answers to the riddle epitomised by the picture on the previous page remain unsatisfying, compared to the explanations that I am accustomed to offering in my books. And indeed, compared to the explanation of egg mimicry. But here’s the best explanation – or series of partial explanations – I can find. We return to the idea of the arms race. In our 1979 paper, John Krebs and I considered ways in which an arms race might end in ‘victory’ for one side (here again, the quotation marks are strongly advised). We identified two principles, the ‘Life Dinner’ and the ‘Rare Enemy’ principle. These are closely related, maybe just different aspects of the same thing.
In one of Aesop’s Fables, a hound was pursuing a hare, got tired and gave up. Taunted for his lack of stamina, the hound replied, ‘It’s all very well for you to laugh, but we had not the same stake at hazard. He was running for his life, but I was only running for my dinner.’
As in military arms races, predators and prey must balance design improvements and resources against economic costs. The more they put into servicing the arms race – muscles, lungs, heart, the machinery of speed and endurance – the less is available for other aspects of life such as making eggs or milk, building up fat reserves for the winter etc. In the language of Darwinism, Aesop’s hares have been subject to stronger selection to invest resources into the arms race than the hounds. There is an asymmetry in the cost of failure – loss of life versus mere loss of dinner. The failed predator lives to pursue another prey. The failed prey has fled its last pursuer. But now, notice how we can say the same thing more piercingly in the language of the genetic book of the dead. The predator’s genes can look back on ancestors many of whom were outrun by prey. But not one of the prey’s ancestors was outrun by a predator. At least not before it had passed on its genes. Plenty of predator genes can look back on ancestors who failed to outrun prey. Not a single prey gene can look back on ancestors who had lost a race against a predator.
Apply the Life Dinner Principle to the cuckoo nestling and its host. The cuckoo nestling can look back on an unbroken line of ancestors, literally not a single one of whom was outwitted by a discriminating host. If it had been, it would not have become an ancestor. Cuckoo genes for failing to fool hosts are never passed on. But genes that lead foster parents to fail to notice cuckoos? Plenty of hosts who were fooled by cuckoos could live to breed again. Genetic tendencies among hosts to be fooled by cuckoos can be passed on. Genetic tendencies among cuckoos to fail to fool hosts are never passed on. It’s the Life Dinner Principle in operation.
Moreover, the host can look back on ancestors many of whom may never have met a cuckoo in their lives. In Nick Davies and Michael Brooke’s long-running study on Wicken Fen, only 5 to 10 per cent of reed warbler nests were parasitised by cuckoos. And this brings us to the Rare Enemy Effect. Cuckoos are comparatively rare. Most reed warblers, wagtails, pipits, dunnocks, etc. probably get through their lives and successfully reproduce without ever encountering a cuckoo. They may look back on many ancestors who never encountered a cuckoo in their lives. But every single cuckoo looks back at an unbroken line of ancestors who successfully fooled a host into feeding them. Asymmetries of this kind could favour ‘victory’ such that even a monstrous cuckoo nestling gets away with fooling its diminutive foster parent. The selection pressure to outwit cuckoos is weak compared to the selection pressure on cuckoos to do the outwitting.
Another parable with an Aesopian flavour is the fable of the boiled frog. A frog dropped into very hot water might do anything in its power to jump out. But a frog in cold water that is slowly heated up does not notice until it is too late. When the baby cuckoo first hatches, the deceiver is indistinguishable from the real thing. As it gradually grows, there is no one day when it suddenly becomes obvious that it’s a fake. Just as there’s never a day when a baby becomes a child; or a child a teenager; or a middle-aged man old. Every day, it looks much the same as the day before. Perhaps this helps the outwitting. Note that the boiled frog effect doesn’t apply to eggs. A cuckoo egg suddenly appears in the nest. It doesn’t gradually become more and more imposterish like a cuckoo nestling.
In another pair of papers already mentioned, Krebs and I proposed that animal communication in general can be seen as manipulation. I discussed this in Chapter 7 in connection with nightingale song bewitching John Keats. Birdsong is known to cause female gonads to swell. This is an example of what we called manipulation. It will not always be to the female’s advantage to submit to it. There will be an arms race between salesmanship and sales-resistance, each side escalating in response to the other. What tricks of salesmanship might the cuckoo nestling employ, in response to the sales-resistance of the host? They’d need to be pretty powerful to outweigh the eventually incongruous mismatch in size between foster parent and cuckoo nestling. But that’s no argument against their existence.
All nestlings open their gapes wide and squawk their appeals for food. If you’re a baby reed warbler, say, the louder you cry, the more likely you are to persuade your parent to drop food into your gape rather than a sibling’s (and there is indeed good Darwinian reason for competition among siblings, even real gene-sharing siblings). On the other hand, loud vocalisation costs vital energy. This applies to baby birds as much as to adults. In one study of wrens at Oxford, the researcher allowed himself to speculate that a male literally sang himself to death. The calling rate and loudness of a baby reed warbler will normally be regulated to an optimum level: enough to compete with siblings, but not so much as to overtax itself or attract predators. The oversized baby cuckoo needs as much food as four young reed warblers. It urges the foster parent on by sounding like a clutch of reed warbler chicks rather than just one very loud reed warbler chick.
Among the ingenious field experiments done by Nick Davies, he and his colleague Rebecca Kilner put a blackbird nestling in a reed warbler nest. The young blackbird was about the same size as a cuckoo nestling. The reed warblers fed it, but at a lower rate than they would normally feed a baby cuckoo. Then the experimenters played their masterstroke: a sound recording of a baby cuckoo piped through a little loudspeaker next to the nest, switched on whenever the baby blackbird was seen to beg. Now the reed warbler adults upped the rate with which they fed the blackbird chick, to a rate appropriate to a baby cuckoo – the same rate as for a clutch of baby reed warblers. And indeed, a recording of four baby reed warblers crying had the same effect. It would seem that baby cuckoo squawks have evolved to become a super-stimulus. Super-stimuli are well attested in experiments on bird behaviour. My old maestro Niko Tinbergen reported that oystercatchers, offered a choice, will preferentially attempt to incubate a dummy egg eight times the volume of their own egg. It’s called a supernormal stimulus. Something like this is what we’d expect as the culmination of an evolutionary arms race, with escalating salesmanship on the cuckoo’s side keeping pace with escalating sales-resistance on the part of the foster parents.
How about a visual equivalent of such a super-stimulus? The open beak of all nestlings is conspicuous, often bright yellow, orange, or red. Doubtless such bright coloration persuades the parents to drop food in, the brighter the gape the greater the chance of their favouring this gape rather than a sibling’s. Reed warbler chicks have a yellow gape. Davies and colleagues found that reed warbler parents gauge their food-fetching efforts according to the total area of yellowness gaping at them in the nest, and also to the rate of begging cries. Cuckoo chicks have a red gape. Is this, perhaps, a stronger stimulus than yellow? An experiment with painted gapes failed to support the hypothesis. Is the cuckoo gape, then, larger than a reed warbler chick’s gape? Yes, cuckoo chicks have a bigger gape than any one reed warbler chick. But its area is not equal to the sum of four reed warbler chicks – perhaps closer to two. Cuckoo chicks use sound to compensate for this, and by two weeks of age a cuckoo chick sounds like a clutch of reed warbler chicks. The combination of a somewhat bigger gape than one reed warbler chick’s, together with supernormal begging cries, is just enough to persuade the adult reed warblers to pump into the cuckoo chick as much food as they would normally bring to a whole clutch of their own chicks. Once again, we could see the supernormal begging call as the end product of an escalating arms race between salesmanship and sales-resistance.
A cardinal feeding a goldfish
That birds are susceptible to large gapes – even the alien gape of a fish – is shown by the well-attested observation of a cardinal (an American bird) repeatedly dropping food into the open mouth of a goldfish. We view the scene through human eyes and think, how absurd, how could a bird be so stupid? But the example of the oystercatcher sitting on the giant egg should warn us that human eyes are precisely what we should not trust. We have no right to be sarcastic. Birds are not little humans, cognitively aware of what they are doing and why they are doing it. And after all, a human male can be sexually aroused by a supernormal caricature of a female, even though he is well aware that it is a drawing on two-dimensional paper, with unnaturally exaggerated features, and a fraction of normal size. The baby cuckoo has no idea what it is doing when it tosses eggs out of the nest. Think of it as a programmed automaton. The oystercatcher does not know why it sits on a giant egg. Think of it as a pre-programmed incubation machine. And in the same way, think of a parent bird as a robot mother, programmed to drop food into wide-open gapes, however ridiculous it may seem to us when the gape belongs to a fish. Or to the giant imposter who is a nestling cuckoo.
If cuckoo nestlings have a supernormal gape, mimicking two ordinary chicks, there’s an Asian cuckoo, Horsfield’s hawk cuckoo, Cuculus fugax, that goes one better. It has the visual equivalent of a clutch of gapes. In addition to its yellow gape, it has a pair of dummy gapes: a patch of bare skin on each wing, the same yellow colour as the real gape. It waves the wing patches about, usually one at a time, next to the real gape. The foster parent (a species of blue robin was the host in this Japanese study by Dr Keita Tanaka) is stimulated by the double whammy of gape plus patch. Dr Tanaka has kindly sent me several photographs plus some amazing film footage. As soon as the foster parent flies in, the cuckoo chick dramatically raises its right wing and waves it about. The gesture reminds me of a swordsman raising his shield to intercept an attack. But this analogy has it exactly wrong. The point is not to repel but to attract. One film even shows the robin vigorously stuffing food up against the yellow patch on the upheld right wing, before turning and shoving it into the wide-open gape instead. The Japanese researchers ingeniously blacked out the wing patch, and this reduced the feeding rate by the robins. There’s a similar story for another brood parasite, the whistling hawk cuckoo, Hierococcyx nisicolor, in China. Like the Horsfield’s hawk cuckoo, the nestlings have yellow wing patches that they display in the same way, to fool foster parents.
So much for cuckoos, not deplorable because a true wonder of nature and natural selection. Now, let’s see what else we can do with the notion of genes looking over their shoulder.
Horsfield’s hawk cuckoo with fake gape on wing
11 More Glances in the Rear-View Mirror
Where once they would have talked of the good of the species, nowadays essentially all serious biologists studying animal behaviour in the wild have adopted what I am calling the gene’s-eye view. Whatever the animal is doing, the question these modern workers ask is, ‘How does the behaviour benefit the self-interested genes that programmed it?’ David Haig, now at Harvard University, is one of those pushing this way of thinking towards the limit, illuminating a great diversity of topics, including some important ones that doctors should care about, such as problems of pregnancy.
Among other things, Haig noticed a lovely example of genes looking backwards – actually at the immediate past generation. There’s a phenomenon called genomic imprinting. A gene can ‘know’ (by a chemical marker) whether it came from the individual’s father or mother. As you can imagine, this radically changes the ‘strategic calculations’ whereby a gene looks after its own self-interest. Haig shows how genomic imprinting changes how a gene views kin. Normally, a gene for kin altruism should regard a half-sibling as equivalent to a nephew or niece – half the value of a full sib or offspring. But if the altruistic gene ‘knows’ it came from the mother and not the father, it should see a maternal half-sibling as equal to its own offspring, or to a normal full sibling. The other way round if it ‘knows’ it came from the father. It should then see the maternal half-sibling as equivalent to an unrelated individual. Genomic imprinting opens up a whole lot of ways in which genes within an individual can come into conflict with one another, the topic of Burt and Trivers’ book Genes in Conflict. Haig goes so far as to blame warring genes for the familiar psychological sensation of being pulled in two directions at once, as in short-term gratification versus longer-term benefit. Genomic imprinting provides a stark example of how a gene might look in the ‘rear-view mirror’. Other examples constitute the topics of this chapter.
A gene on a mammalian Y-chromosome ‘looks back’ at an immensely long string of ancestral male bodies and not a single female one, probably as far back as the dawn of mammals if not further. Our mammal Y-chromosome has been swimming in testosterone for perhaps 200 million years. But if Y-chromosomes look back at only male bodies, what about X-chromosomes? If you are a gene on an X-chromosome, you might come from the animal’s father, but you are twice as likely to come from its mother. Two-thirds of your ancestral history has been in female bodies, one-third in male bodies. If you are a gene on a chromosome other than a sex chromosome, an autosome, half your ancestral history was in female bodies, half in male bodies. We should expect many autosomal genes to have sex-limited effects, programmed with an IF statement: one effect whenever they find themselves in a male body, a different effect when in a female body.
But when any gene looks back at the male bodies that it has inhabited, what it sees will not be a random sample of male bodies but a restricted sub-set. This is because the average male is often denied the Darwinian privilege of reproduction. A minority of males monopolises the mating opportunities. Most females, on the other hand, enjoy close to the average reproductive success. Red deer stags with large antlers prevail in fights over access to females. So when a red deer gene looks back at its male ancestors, it will see the minority of male bodies that are topped by abnormally large antlers.
Even more extreme is the asymmetry shown by seals, especially Mirounga, the elephant seal. There are two species: the southern elephant seal, which I have seen, close enough to touch (though I would not), on the remote island of South Georgia, and the northern elephant seal, which Burney Le Boeuf has thoroughly studied on the Pacific beaches of California. Like many mammals, elephant seals have harem-based societies but they carry it to an extreme. Successful males, ‘beachmasters’, are gigantic: up to 4 metres long and weighing 2 tonnes. Females are relatively small and are gathered into harems, which may typically number as many as fifty ‘belonging to’, and vigorously defended by, a single dominant male. Most of the males in the population have no harem, and either never reproduce or bide their time hoping to sneak an occasional copulation, as well as aspiring eventually to get big and strong enough to displace a beachmaster. In one report from Le Boeuf’s long-term California study of northern elephant seals, only eight males inseminated an astonishing 348 females. One male inseminated 121 females, while the great majority of males had no reproductive success at all. An elephant seal gene on a Y-chromosome looks back at, not just a long sequence of male bodies, but specifically at the overgrown, blubbery, belching, bloated bodies of a tiny minority of dominant, harem-holding beachmasters: highly aggressive males, over-endowed with testosterone and with the dangling trunks used as living trombones to resonate roars that intimidate other males. On the other hand, an elephant seal gene will look back at a succession of female bodies that are close to the average.
Do you find something puzzling about the fact that only a small minority of males does almost all the fathering? Isn’t it terribly wasteful? Think of all those bachelor males, consuming a fat slice of the food resources available to the species, yet never reproducing. A ‘top-down’ economic planner with species welfare in mind would protest that most of those males shouldn’t be there. Why doesn’t the species evolve a skewed sex ratio such that only a few males are born: just enough males to service the females, the same number of males as would normally hold harems? They wouldn’t have to fight each other, they’d all get a harem as a matter of automatic entitlement, just for being male. Wouldn’t a species with such an economically sensible, planned economy prevail over the present, wildly uneconomical, strife-ridden species? Wouldn’t the planned economy species win out in natural selection?
Sexual inequality on the beach
Yes, if natural selection chose between species. But, contrary to a widespread misunderstanding, it doesn’t. Natural selection chooses between genes, by virtue of their influence on individuals. And that makes all the difference. If the sensible planned economy were to come about by Darwinian means, it would have to be through the natural selection of genes controlling the sex ratio. This is not impossible. A gene could bias the number of X sperms versus Y sperms produced by males. Or it could favour selective abortion of some male foetuses. Or it could favour starving some baby sons to death and keeping just a favoured few. Never mind how it does it, just call this hypothetical gene the Planned Economy Gene, pegged to top-down common sense.
Imagine a planned economy population where most of the individuals are female, say one male for every ten females. This is the kind of population our sensible economist would expect to see. It is economically sensible because food is not wasted on males who are never going to reproduce. Now imagine a mutant gene arising, a mutation that biases individuals towards having sons. Will this male-favouring gene spread through the population? Alas for the planned economy, it certainly will. In the planned economy, females outnumber males ten to one, so a typical male can expect ten times as many descendants as a typical female. It’s a bonanza for males. The son-biased mutant gene will spread rapidly through the population. And the males will have good reason to fight. It’s the flip side of our observation that our hypothetical gene looks back at a successful minority of male bodies, not at an average sample of male bodies.
Will the population sex ratio swing right round to the opposite extreme and become male-biased? No, natural selection will stabilise the sex ratio we actually see, a 50/50 sex ratio (but see the important reservation below) with a minority of harem-holding males and a majority of frustrated bachelors. Here’s why. If you have a son, there’s a good chance he’ll end up a disconsolate bachelor who’ll give you no grandchildren. But if your son does end up a harem-holder, you’ve hit the jackpot where grandchildren are concerned. The expected reproductive success of a son, averaged over his slim chance of the jackpot plus the much greater chance of bachelor misery, equals the expected average reproductive success of a daughter. Equal sex ratio genes prevail, even though the society they create is so horribly uneconomical. Sensible as it sounds, the ‘planned economy’ cannot be favoured by natural selection. In this respect at least, natural selection is not a ‘sensible’ economist.
I said that selection would stabilise the sex ratio at 50/50 but I added a cautionary reservation. There are various reasons for that caution, and they are important. Here’s one of them. Suppose it costs twice as much to rear a son as to rear a daughter. To equip a son to fight off rivals and win a harem, he must be big. Being big doesn’t come free. It costs food. If a mother seal must suckle a son for longer than a daughter, if a son costs twice as much as a daughter to rear, the ‘choice’ facing the mother is not ‘Shall I have a son or a daughter’ but ‘Shall I have a son or two daughters?’ The general principle, first clearly understood by RA Fisher, is that the sex ratio stabilised by natural selection is 50/50 measured in economic expenditure on daughters versus economic expenditure on sons. That will amount to 50/50 in numbers of male and female bodies, only if the cost of making sons and daughters is the same. Fisher’s principle balances what he called parental expenditure on sons versus daughters. This may cash out in the form of equal numbers of males and females in the population, but only if sons and daughters are equally costly to rear. There are other complications, some pointed out by WD Hamilton, but I won’t stay to deal with them.
Elephant seals are an extreme example of a principle that typifies many mammal species. Females tend to have nearly the same reproductive success as each other, close to the population average, while a minority of males enjoys a disproportionate monopoly of reproduction. In statistical language, mean reproductive success of males and females is equal, but males tend to have a higher variance in reproductive success. And, to return to the title of this chapter, the ancestral females that genes ‘look back on’ will be close to the average. But they’ll look back on an ancestral history dominated by a minority of males: that minority endowed with whatever it takes in the species concerned – large antlers, fearsome canine teeth, sheer bodily bulk, courage, or whatever it might be.
‘Courage’ can be given a more precise meaning. Any animal must balance the short-term value of reproducing now against its own long-term survival to reproduce in the future. A brutal fight against a rival male may end in victory and a harem. But it may end in death, or serious injury which presages death. Courage is at a premium. Risking death is worthwhile because the stakes for a male are so high: a huge number of pups to his name if he wins, zero and perhaps death if he loses. A female seal would give higher priority to surviving to reproduce next year. She only has one pup in a year, so she’ll maximise her reproductive success by surviving herself. Natural selection would favour females who are more risk-averse than males; would favour males who are more courageous or foolhardy. Males are biased towards a high-stakes high-risk strategy. This is probably why males tend to die younger. Even if they’re not killed in battle, their whole physiology is skewed towards living to the full while young, even at the expense of living on at all when old.
A complication is that, in some species, including elephant seals, subordinate males sneak surreptitious matings at the risk of punishment from dominant males. They may adopt a particular strategy known as the ‘sneaky male’ strategy. This means that as a Y-chromosome looks back at its history, it will see mostly a river of dominant harem-holders but also a side rivulet, that of the sneaky males. And now, a change of topic.
As will be apparent by now, my late colleague WD Hamilton had a restless and highly original curiosity, which led him to solve many outstanding riddles in evolutionary theory, problems that lesser intellects never even recognised as problems. A naturalist from boyhood, he noticed that many insect species come in two distinct types which could be named ‘dispersers’ and ‘stay-at-homes’. Dispersers typically have wings. ‘Stay-at-homes’ often don’t. It’s surprising how many species of insects have both winged and wingless members, seemingly in balanced proportion. If you like human parallels, think of human families in which one brother comfortably inherits the farm while the other brother emigrates to the far side of the world in search of an improbable fortune. In the case of plants, dandelion seeds with their fluffy parachutes are ‘winged’ dispersers, while other members of the daisy family have, to quote Hamilton, ‘a mixture of winged and wingless within a single flower head’.
To stolid common sense, it seems intuitively obvious that if parents live in a good place (and they probably do live in a good place, or they wouldn’t have succeeded in becoming parents), the best strategy for an offspring must be to stay in the same good place. ‘Stay at home and mind the family farm’ would seem to be the watchword, and that was the conventional wisdom among most evolutionary theorists before Bill Hamilton. Bill suspected, by contrast, that selection would favour a balance between stay-at-homes and dispersers, the point of balance varying from species to species. He enlisted the help of his mathematical colleague Robert May, and together they developed mathematical models that supported his intuition.
My own, less mathematical way to express Bill’s intuition is in terms of the gene’s-eye view of the past. No matter how favourable the ‘family farm’ – the environment in which parents have flourished – it is sooner or later going to be subject to a catastrophe: a forest fire perhaps, or a disastrous flood or drought. So, as a gene looks back at the history of ‘the family farm’, the parental, grandparental, and great grandparental generations may indeed have flourished there. The success story might go back an unbroken ten or even twenty generations. But eventually, if it looks far enough back into the past, the stay-at-home gene will eventually hit one of those catastrophes.
The disperser gene may look back on the recent past as one of comparative failure: life on the family farm was milk and honey. But if we look back sufficiently far, we come to a generation where only the disperser gene, the gene for wild wanderlust, made it through. There’s also the anthropomorphic point that wanderlust occasionally strikes gold.
Naked mole rat
I perhaps went too far when in 1989 I published a speculation about naked mole rats, but it serves to dramatise the point. Naked mole rats are small, spectacularly ugly (by human aesthetics) African mammals, who live underground. They are famous among biologists as the nearest mammalian approach to social insects: ants and termites. They live in large colonies of as many as 100 individuals in which only one female, the ‘queen’, normally reproduces, and she is fecund enough to compensate for the near sterility of all the other females, who function as ‘workers’. A colony can extend through a huge network of 2 or 3 miles of burrows, gathering underground tubers as food.
This much has become lore among biologists intrigued by the obvious similarity to social insects. However, one discrepancy always worried me. Although the ants and termites that we ordinarily see are wingless, sterile workers, their underground nests periodically erupt in a boiling mass of winged reproductive individuals of both sexes. These fly up to mate, after which the newly fertilised young queens settle down, lose their wings (in many cases even biting them off), dig a hole, and attempt to found a new underground nest with the aid of sterile, wingless worker daughters (and sons in the case of termites). The winged castes are Hamilton’s dispersers, and they are an essential part – indeed, the essential part – of the biology of social insects. You could say they are what the whole social insect enterprise is all about. Why don’t naked mole rats have an equivalent? Their lack of a dispersal phase is something approaching a scandal!
Not literally winged dispersers! Even I am not foolhardy enough to predict rodents with wings. But I did wonder, and still do, whether there might be a dispersal phase that nobody has spotted yet. In 1989 I wrote: ‘Is it conceivable that some already known hairy rodent, running energetically above ground and hitherto classified as a different species, might turn out to be the lost caste of the naked mole rat?’ My idea for a hitherto unrecognised dispersal caste may not have much going for it, but it is at least testable, a virtue that scientists value highly. The genome of the naked mole rat has been sequenced. If my hypothetical dispersal phase were ever discovered, some hairy mole rats should turn out to have the same genes.
I admitted the implausibility of my suggestion. How could such a hypothetical creature have been overlooked by biologists? However, I went on to make a comparison with locusts. Locusts are the terrifying ‘wanderlust’ phase of harmless ‘stay-at-home’ grasshoppers. They look different from grasshoppers and behave very differently. They are the very same grasshoppers but (oh, in a moment) they change. The genes of a harmless grasshopper have the capacity, when the conditions are right, to change (change utterly, and a terrible beauty is born). The devastating effects are all too well known. My point is that locust plagues only occasionally happen. It just takes the right conditions. Perhaps the dispersal phase of the naked mole rat has yet to erupt during the decades since biologists have been around to study the species? No wonder it has never yet been seen. Perhaps it would take only a crafty hormone injection … and a naked mole rat could become its own hairy, scurrying (though not, I suppose, winged) dispersal phase.
Another change of topic before we leave the backwards gene’s-eye view. There are two ways in which we can look back at a family tree. Conventional pedigrees trace ancestry via individuals. Who begat whom? Which individual was born of which mother? The most recent individual ancestor shared by the late Queen Elizabeth II and her husband Prince Philip was Queen Victoria. But you can also trace the ancestry of a particular gene, and you will have guessed that this is the alternative manner of tale I want to tell here. Genes, like individuals, have parent genes and offspring genes. Genes, as well as individuals, have pedigrees, family trees. But there is a significant difference between a ‘people tree’ and a ‘gene tree’. An individual person has two parents, four grandparents, eight great grandparents, etc. So a people tree is a vast ramification as you look backwards in time. Any attempt to draw it out completely will soon get out of hand. The best way to visualise it is not on paper but zooming around a computer screen. Not so the gene tree. A gene has only one parent, one grandparent, one great grandparent, etc. A gene tree is therefore a simple linear array streaking back in time, whereas a people tree bifurcates its way unmanageably into the past. This is not so when you look forwards in time, by the way. A gene can have many offspring but only ever one parent. Looking forwards, gene trees branch and branch. But this chapter is all about looking backwards.
A particular sub-lethal gene, haemophilia, has plagued the royal families of Europe ever since the early nineteenth century. The gene tree of royal haemophilia is simple and fits the page comfortably. The equivalent people tree would want several square metres of paper to be legible. The royal haemophilia gene can be traced back to a particular individual ancestor, Queen Victoria, one of whose two X-chromosomes bore the gene. The mutation occurred, to quote Steve Jones’s mordant phrase, ‘in the august testicles’ of her father, Edward, Duke of Kent. One of Victoria’s four sons, Prince Leopold, suffered from haemophilia. The other sons, including Edward VII and his descendants such as our present monarch, King Charles III, beat the odds and were lucky to escape. Leopold survived to the age of thirty, long enough to have a daughter, Princess Alice of Albany, who inevitably carried the gene on one of her X-chromosomes. Her son Prince Rupert of Teck realised his 50 per cent probability of being afflicted and died young.
Royal haemophilia
Of Victoria’s five daughters, three (at least) inherited the gene. Princess Alice of Hesse passed it on to her son, Prince Friedrich, who died in infancy, and to two daughters, Irene and Alexandra, who passed it on to three haemophiliac grandsons of Alice, including the Tsarevich Alexey of Russia. Irene married her first cousin Henry, a common practice among royals and generally not a good idea because of inbreeding depression. But inbreeding depression was not responsible for the fact that two of their sons, Waldemar and Heinrich, suffered from haemophilia: they got it on their X-chromosome from their mother, and she’d have been equally likely to pass it on, whomever she married, cousin or not (unless the cousin was himself haemophiliac, in which case 50 per cent of her daughters would actually suffer from the disease itself). Another of Victoria’s daughters, Princess Beatrice bequeathed the gene to her daughter the Queen of Spain, and on into the Spanish Royal Family, to the resentment, I gather, of the Spanish.
Tracing back the gene tree of the royal haemophilia gene, all lines coalesce in Victoria. And indeed, there is a flourishing branch of mathematical genetic theory called Coalescent Theory in which you look back at the history of a genetic variant in a population and trace the most recent common ancestor of that gene – the coalescent gene upon which all lines converge as you look back. Forget about individuals, look through the skin to the genes within, and you can trace two copies of a particular gene back in time until you hit the ancestor in whom they coalesce. That coalescence point is the ancestral individual in which the gene itself divided into two copies, which then went their separate ways in two siblings and eventually two lines of descendants. If you make purifying assumptions like random mating, no natural selection, and everybody has two children, the coalescent tree has an expected form that mathematicians can calculate in theory. In reality, of course, those assumptions are violated, and that’s when it becomes interesting. Royal families, for example, typically violate the assumption of random mating. Protocol and political expediency constrain them to marry each other.
Coalescent theory is an important part of modern population genetics, and very relevant to this chapter on the backwards gene’s-eye view, but the mathematics is outside my scope here. I will discuss one intriguing example: a particular study of one man’s genome – as it happens, my genome, although that isn’t why I find it intriguing. It is a remarkable fact that you can make powerful inferences about the demographic history of an entire population using the genome of just a single individual. For a rather odd reason, I was one of the earliest people in Britain to have their entire genome (as opposed to the relatively small sample done by the likes of ‘23-and-Me’) sequenced. I handed the data disc over to my colleague Dr Yan Wong, and he included a clever analysis of it in the book that we co-authored, The Ancestor’s Tale (2016). It’s rather tricky to explain, but I’ll do my best.
In every cell of my body swim twenty-three chromosomes inherited intact from my father and twenty-three from my mother. Every (autosomal) paternal gene has an exact opposite number (allele) on the corresponding maternal chromosome, but my father John’s chromosomes and my mother Jean’s chromosomes float intact and aloof from each other in all my cells. Now, here’s where it gets tricky. Take a particular gene on a John chromosome and allow it to look back at its ancestral history. Now take its opposite number (‘allele’) on the equivalent Jean chromosome, and allow it to look back in the same way. It’s the same principle as tracing the royal haemophilia gene back to Victoria. But, in this case, it is not haemophilia that is being traced, we’re looking a lot further back, and we have no hope of identifying a named individual like Victoria. We could do it with any pair of alleles, one on a John chromosome and the other on a Jean chromosome. And not just one such pair but (a sample of the) many.
Sooner or later, each gene pair, as they look back, is bound to converge on a particular individual in whom a gene once split to form the ancestor of the John gene and the ancestor of the Jean gene. I really do mean a particular individual ancestor who lived at a particular time and in a particular place. This individual had two children, one of whom was John’s ancestor and the other Jean’s ancestor. But we’re talking about a different ancestral individual – different time and place – for each Jean/John gene pair. For each gene pair, there must have been two siblings, one carrying the ancestral Jean gene and the other the ancestral John gene.
There are many overlapping people-tree routes that trace my father and my mother back to different shared ancestors. But for each of my John genes there is only one path linking it to the shared ancestor of my corresponding Jean gene. Gene trees are not the same as people trees. Each gene pair coalesces in a particular ancestor, at a particular moment in the past. You can let each pair of my genes look back, and you can find a different coalescence point in each case. You can’t literally identify the exact coalescence point for any given gene pair. But what you can do, using the mathematics of coalescent theory, is estimate when it occurred. When Dr Wong did this with my genome, he found that a large majority coalesced somewhere around 60,000 years ago, say 50,000 to 70,000.
And how should this concordance be interpreted? It means that my ancestors suffered a population bottleneck around that time. Very likely, yours did too. As my John genes and my Jean genes look back at their history, during most of those millennia they see a picture of outbreeding. But somewhere around 60,000 years into the past, the effective population size narrowed to a bottleneck. When the population is smaller, the Jean and John lineages are more likely to find themselves in a shared ancestor, simply by chance. That is why my gene pairs tend to coalesce around that time. Indeed, the coalescence data from my genome, on its own, making use of no other data, can be translated into the above graph of effective population size plotted against time. It is presumably typical for Europeans. The faint grey line shows the equivalent for an individual Nigerian, whose ancestors, it would seem, were not subject to the same bottleneck. I confess to an obscure satisfaction that, of the two co-authors of a book, one was able to use the genome of the other to make a quantitative estimate of prehistoric demography affecting not just one individual but millions.
What else can genes tell us as they look back at their history? Zoologists are accustomed to drawing family trees of animals, and calculating which species are close cousins of other species, and which distant. Among ape species, for example, chimpanzees and bonobos are our closest living relatives, and those two species are exactly equally close to us. They are equally close because they share an ancestor with each other some 3 million years ago, and that ancestor shares an ancestor with us about 6 million years ago (see below). Gorillas are the outgroup, a more distant relative of the rest of us African apes. The ancestor we share with gorillas lived longer ago, perhaps 8 or 9 million years.
GORILLA
CHIMP
BONBO
HUMAN
On the previous page is the conventional way to draw a family tree, an organism-based family tree. But we can also draw a family tree from the point of view of a gene, looking back at its own history. The organism tree is unequivocal. Chimps and bonobos are close cousins of each other, and we are their closest relatives apart from each other. But while that is indeed a fact from the point of view of the whole organism, it is not necessarily the case when it is genes that look in the rear-view mirror. True, a majority of genes would ‘agree’ with each other and with the ‘people tree’ of the traditional zoologist. Nevertheless, it is perfectly possible that, from the point of view of some particular genes, the family tree could look very different. As on the opposite page, perhaps. The majority of our genes agree with the ‘people tree’. But when the gorilla genome was published in 2012, it turned out that ‘Humans and chimpanzees are genetically closest to each other over most of the genome, but the team found many places where this is not the case. Fifteen per cent of the human genome is closer to the gorilla genome than it is to chimpanzee, and 15 per cent of the chimpanzee genome is closer to the gorilla than human.’ I hope you agree that his kind of conclusion is an interesting product of the ‘backward gene’s-eye view’.
Such an anomaly could occur even within one small family. Two brothers, John and Bill, share the same parents, Enid and Tony, and the same four grandparents: Arthur and Gertrude, the parents of Enid, and Francis and Alice, the parents of Tony. (Sex chromosomes apart) each of the brothers received exactly half his genes from each of their shared parents. That’s because each is the product of exactly one egg from Enid and one sperm from Tony. And each brother received a quarter of his genes from each of the four shared grandparents, but in this case the figure is only approximate. It’s not exactly a quarter. Through the vagaries of chromosomal crossing-over, the sperm from Tony that conceived John could, by chance, have contained mostly Alice’s genes rather than Francis’s. The sperm from Tony that conceived Bill could have contained a preponderance of Francis’s genes rather than Alice’s. The egg from Enid that gave rise to John could have contained mostly Arthur’s genes, while the egg from Enid that gave rise to Bill contained a preponderance of Gertrude’s genes. It’s even theoretically possible (though vanishingly improbable) that John received all his genes from two of his grandparents, and none from the other two. Thus, the gene’s-eye view of closeness of relatedness can differ from the individual’s-eye view. The individual’s-eye view sees all four grandparents as equal contributors.
BONOBO
CHIMP
GORILA
HUMAN
And the same is true of all generations prior to the immediate parental generation. Although you are quite probably descended from William the Conqueror, it is also quite likely that you have inherited not a single gene from him. Biologists tend to follow the historic precedent of tracing ancestry at the level of the whole individual organism: every individual has one father and one mother, and so on back. But the John/Bill, gorilla/chimpanzee comparison of the previous paragraphs will prove, I believe, to be the tip of an iceberg. More and more, we shall see pedigrees being drawn up from the genes’ point of view as opposed to the individual organism’s. An example is the discussion of the prestin gene in Chapter 5. Such a trend is obviously highly congenial to this book, stressing, as it does, the gene’s-eye view.
The last topic I want to deal with in this chapter on the backwards gene’s-eye view is Selective Sweeps. Among the messages from the past that the genes of a living animal whisper to us, if only we could hear them, many tell of ancient natural selection pressures. That, indeed, is what I mean by the genetic book of the dead, but here I am talking about a particular kind of signal from the past, one that geneticists have learned how to read. Present-day genes send statistical ‘signals’ of natural selection pressures. A gene pool that has recently undergone strong selection shows a certain characteristic signature. Natural selection leaves its mark. A Darwinian signature. Here’s how.
Two genes that sit close to one another on a chromosome tend to travel together through the generations. This is because chromosomal crossing over is relatively unlikely to split them: a simple consequence of their proximity to each other. If one gene is strongly favoured by natural selection it will increase in frequency. Of course, but mark the sequel. Genes whose chromosomal position lies close to a positively selected gene will also increase in frequency: they ‘hitch-hike’. This is especially noticeable when the linked genes are neutral – neither good nor bad for survival. When a particular region of a chromosome contains a gene that is under strong selection in its favour, the geneticist notices a diminution in the amount of variation in the population, specifically in the hitch-hiking zone of the affected chromosome. Because of the hitch-hiking, natural selection of one favoured gene ‘sweeps’ away the variation among nearby neutral genes. This ‘selective sweep’ then shows up as a ‘signature’ of selection.
I find the ‘backwards’ way of looking at ancestral history illuminating. But the most important ‘experience’ that a gene can ‘look back on’ is easily overlooked because it hides in plain view. It is the companionship of other genes of the species: other genes with which it has had to share a succession of bodies. I am not talking here about genes being linked close to each other on the same chromosome. I am now talking about shared membership of the same gene pool, and hence of many individual bodies. This companionship is the topic of the next chapter.
12 Good Companions, Bad Companions
The previous chapter could be expanded with an indefinite number of examples of the backward gene’s-eye view. Genes look back on a series of environments variously characterised by trees, soil, predators, prey, parasites, food plants, water holes, etc. But the external environment is only part of the story. It leaves out the most important kind of ‘experience’ of a gene. Far more important is the experience of rubbing shoulders with all the other genes in a long succession of bodies: partners through dynasties of mutual collaboration in the subtle arts of building bodies. That is the central point of this chapter.
The genes within any one gene pool are travelling bands of good companions, journeying together, and cooperating with each other down the generations. Genes in other gene pools, gene pools belonging to other species, constitute parallel bands of travelling companions. These bands do not include the genes of other species. That is precisely how biologists like to define a species (although the definition sometimes blurs in practice, especially when new species are being born).
Sexual reproduction validates the very notion of a species, more precisely the notion of a gene pool: a pool of genes like a stirred pool of water. The gene pool is thoroughly stirred in every generation by sexual reproduction, but it doesn’t mix with any other such pool – pools belonging to other species. Children resemble their parents but, because the gene pool is stirred, they resemble them only slightly more than they resemble any random member of the species – and much more than they resemble a random member of another species. The gene pool of each species sloshes about in a watertight compartment of its own, isolated from all others.
As I said, that is part of the very definition of a ‘species’, at least the most widely adopted definition, the one codified by that lofty patriarch among evolutionists, Ernst Mayr (1904–2005):
Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups.
Fossils, being dead to the possibility of actually interbreeding – beyond breeding at all – force a retreat to Mayr’s ‘potentially’. When we say that Homo erectus was a separate species, distinct from modern Homo sapiens, the Mayr definition would be interpreted as meaning, ‘If a time machine enabled us to meet Homo erectus, we would be incapable of interbreeding with them.’ A niggling difficulty arises over ‘incapable’. There are species that can be persuaded to interbreed in captivity but would not choose to do so in the wild. Chapter 9’s example of the two crickets Teleogryllus oceanicus and commodus is only one of several. Even if we were capable of interbreeding with Homo erectus, say by artificial insemination, would we – or they – choose to do so by the normal, natural means? Never mind, that is a detail that might concern a pernickety taxonomist or philosopher, but we can pass it by.
If, as most anthropologists believe, we descend from Homo erectus, there must have been intermediates during the transitional phase: intermediates that would defy classification. Nobody who has thought it through would suggest that suddenly a sapiens baby was born to proud erectus parents. Every animal ever born throughout evolutionary history would have been classified in the same species as its parents, not only by the interbreeding criterion but by all sensible criteria. That fact – though it troubles some minds – is totally compatible with the fact that Homo sapiens is descended from Homo erectus, those two species being distinct species incapable – let us presume – of breeding with each other. It’s also compatible with the fact that you are descended from a lobe-finned fish, with every intermediate along the way being a member of the same species as its parents and its children.
Moreover, when a species splits into two daughter species in the process known as speciation, there is bound to be an interregnum when the two are still capable of interbreeding. The split originates accidentally, imposed perhaps by a geographic barrier such as a mountain range or a river or stretch of sea. It is probable that chimpanzees and bonobos started to go their separate evolutionary ways when two sub-populations found themselves on opposite sides of the Congo river. The two populations were physically prevented from interbreeding – the flow of genes was halted by the flow of water between them. For a while, they could potentially interbreed, and maybe occasionally did so when an individual inadvertently crossed the river on a floating log. But the geographically imposed lack of gene flow freed them to evolve in separate directions. Those different directions could have been guided by natural selection, or unguided in a process of random drift. It doesn’t matter, the point is that the compatibility between their genes gradually declined until a stage was reached when, even if they should chance to meet, they could no longer interbreed in actuality. The initial geographic barrier doesn’t necessarily come about through an environmental change like an earthquake diverting a river. Geography can stay the same while a pregnant female, for instance, gets accidentally washed ashore on a deserted island. Or the other side of a river.
But why, in any case, do the genes of two separated populations tend to become incompatible as companions, thereby preventing interbreeding? One reason is that the two sets of chromosomes need to pair off in the process of meiosis, when gametes are made. If they become sufficiently different, say on opposite sides of a barrier, hybrids, if any, would be unable to make gametes. They might live, but could not reproduce. Another reason – back to the central point of this chapter – is that genes, on either side of the barrier, are naturally selected to cooperate with other genes on the same side, but not the other. After enough time has elapsed in physically enforced separation, two gene pools become so incompatible that interbreeding becomes impossible even if the physical barrier is removed. Chimpanzees and bonobos haven’t quite reached that stage. Hybrids can be born in captivity.
There doesn’t have to be a distinct barrier, like a river, for geographically based speciation to occur. A mouse in Madrid never meets a mouse in Vladivostok but there could well be continuous local gene flow across the 12,000-kilometre gap between them. Given enough time, their descendants could diverge genetically until they could no longer interbreed even if they should somehow contrive to meet. Speciation would have occurred, the barrier being nothing more than sheer distance rather than an unswimmable river or sea, or an impassable desert or mountain range, and despite continuous gene flow locally across the entire range. We have here the spatial equivalent of the temporal continuum between Homo erectus and Homo sapiens. In both cases the extremes never meet. Yet in both cases there can be an unbroken chain of intermediates happily breeding all the way across the range: range in space for the example of the mice; range in time for the example of erectus and sapiens.
Occasionally, the chain of intermediates wraps around in a circle, bites itself in the tail, and we have a so-called ‘ring species’. Salamanders of the genus Ensatina live all around the four edges of California’s Central Valley but don’t cross the valley. If you start sampling at the southern end of the valley and work your way up the west side to the north, go eastwards across the north end of the valley, then down the eastern side and back around to your starting point, you notice a fascinating thing. The salamanders all along your route around the edge of the valley can interbreed with their neighbours. Yet they gradually change as you go around, and when you arrive back at your starting point, the ‘last’ species of the ring cannot interbreed with the ‘first’. A ring species is a rare case where you can see laid out in the spatial dimension the kind of evolutionary change that you could see along the time dimension if only you lived long enough.
Such considerations render pointless all heated arguments about whether or not closely related animals, living or fossil, belong to the same species. It is a necessary consequence of evolution that there must be, or must have been, intermediates that you cannot forcibly assign to either species. It would be worrying if it were otherwise. But of course most species in existence are clearly distinct from most other species by any criterion, because of the long time that has elapsed since their ancestors diverged. As for the grey areas where potential interbreeding is even an issue, and where species definition is problematical, this chapter will not treat them further.
Where external environments are concerned, the genes of a mole speak to us of damp, dark, moist tunnels, of earthy smells, of earthworms and beetle larvae crawling between tangled rootlets and filaments of fungal mycelium and mycorrhizae. The genes of a squirrel have a very different ancestral autobiography, a tale of airy greenery, waving boughs, acorns, nuts, and sunlit glades to be crossed between trees. We could weave a similar list for any species. The point of this chapter, on the other hand, is that the genes’ external ‘experience’ of damp, dark soil, or forest canopy, grassy plains, coral reefs, the deep sea, or whatever it might be, is swamped by the more immediate and salient internal experiencing of other genes in the stirred gene pool. This chapter is about the ‘good companions’ with which the genes have travelled and collaborated, in body after body since earlier times: parting from and re-joining, ever encountering and re-encountering familiar sets of companion genes, collaborating in the difficult arts of building livers and hearts, bones and skin, blood corpuscles and brain cells. The details will be tweaked by ‘external’ pressures: the best heart, kidney, or intestine for a burrowing vermivore is doubtless not the same as the best heart, kidney, or intestine for a tree-climbing nut-lover. But a centrally important quality of a successful gene will be the ability to collaborate with the other genes of the shared gene pool, be it mole, squirrel, hedgehog, whale, or human gene pool.
Every biochemistry lab has on its wall a huge chart of metabolic pathways, a bewildering spaghetti of chemical formulae joined by arrows. Below is a simplified version in which chemicals are represented by blobs rather than having their formulae spelled out. The lines represent chemical pathways between the blobs. This particular diagram refers to the gut bacterium Escherichia coli, but something similar, and just as bewildering, is going on in your cells.
Every one of those hundreds of lines is a chemical reaction performed inside a living cell, and each one is catalysed by an enzyme.
Every enzyme is assembled under the influence of a specific gene (or often two or three genes, because the enzyme molecule may have several ‘domains’ wrapped around each other, each domain being a protein chain). The genes that make these enzymes must cooperate, must be good companion genes in the sense of this chapter.
All mammals have almost exactly the same set of over 200 named bones, connected in the same order, but differing in size and shape. We saw the principle in the crustaceans of Chapter 6. And the same is true of the metabolic pathways diagrammed above. They are almost the same in all animals but different in detail. And, although they may be engaged in joint enterprises that are similar, the cartels of mutually compatible genes will not be compatible with parallel cartels evolving in other lineages: antelope cartels versus lion cartels, say. Antelopes and lions both need metabolic pathways in all their cells, and both need hearts, kidneys, and lungs, but they’ll differ in details appropriate to herbivores versus carnivores. And more obviously so in teeth, intestines, and feet, for reasons we’ve covered already. If they were somehow to mix in the same body, they wouldn’t work well together.
I shall say that two separate gene pools, for instance an impala gene pool and a leopard gene pool, represent two separate ‘syndicates’ of ‘cooperating’ genes. Building a body is an embryological enterprise of immense complexity, involving feats of cooperation between all the genes in the active genome. Different kinds of body require different embryological ‘skills’, perfected over evolutionary time by different suites of mutually compatible genes: compatible with members of their own syndicate but incompatible with other syndicates simultaneously being built in other gene pools. These cooperating cartels are assembled over generations of natural selection. The way it works is that each gene is selected for its compatibility with other genes in the gene pool, and vice versa. So cartels of mutually compatible, cooperating genes build up. It is tempting but misleading to speak of alternative cartels being selected as whole units versus other cartels as whole units. Rather, cartels assemble themselves because each member gene is separately selected for its compatibility with other genes within the cartel, which are themselves being selected at the same time.
Within any one species, genes work together in embryological harmony to produce bodies of the species’ own type. Other cartels in other species’ gene pools self-assemble, and work together to produce different bodies. There will be carnivore cartels, herbivore cartels, burrowing insectivore cartels, river-fishing cartels, tree-climbing, nut-loving cartels, and so on. My main point in this chapter on ‘Good Companions’ is that by far the most important environment that a gene has to master is the collection of other genes in its own gene pool, the collection of other genes that it is likely to meet in successive bodies as the generations go by. Yes, the external ecosystem furnished by predators and prey, parasites and hosts, soil and weather, matters to the survival of a gene in its gene pool. But of more pressing moment is the ecosystem provided by the other genes in the gene pool, the other genes with which each gene is called upon to cooperate in the construction and maintenance of a continuing sequence of bodies. It is an easily dispelled paradox that my first book, The Selfish Gene, could equally well have been called The Cooperative Gene. Indeed, my friend and former student Mark Ridley wrote a fine book with that very title. In his words, which I’d have been pleased to have written myself,
The cooperation between the genes of a body did not just happen. It required special mechanisms for it to evolve, mechanisms that arrange affairs such that each gene is maximally selfish by being maximally cooperative with the other genes in its body.
As inhabitants of today’s technologically advanced world, we are aware of the power of cooperation between huge numbers of specialist experts. SpaceX employs some 10,000 people, cooperating in the joint enterprise of launching massive rockets into space and – even more difficult – bringing them back and gently landing them in a fit state to be re-used. Many different specialists are united in intimate cooperation: engineers, mathematicians, designers, welders, riveters, fitters, turners, computer programmers, crane operators, quality control checkers, 3-D printer operators, software coders, inventory control officers, accountants, lawyers, office workers, personal assistants, middle managers, and many others. Most of the experts in one field have little understanding of what experts in other parts of the enterprise do, or how to do it. Yet the feats that we humans can achieve when thousands of us deploy our complementary skills, in well-oiled collaboration but in ignorance of each other’s role, are staggering.
The human genome project, the James Webb Telescope, the building of a skyscraper or a preposterously oversized cruise ship, these are stunning achievements of cooperation. The Large Hadron Collider at CERN brings together some 10,000 physicists and engineers from more than 100 countries, speaking dozens of languages, working smoothly together to pool their diverse expertise. Yet these huge accomplishments of mass cooperation are more than matched by the nine-month collaborative enterprise of building each one of us in our mother’s womb: a feat of cooperation among billions of cells, belonging to hundreds of cell types (different ‘professions’), orchestrated by about 30,000 intimately cooperating genes, exceeding the personnel count we find in a large human enterprise such as SpaceX. Cooperation is key, in both building a body and building a rocket.
The genes that build a body must cooperate with all the other companions that the sexual lottery throws at them as the generations go by. They must cooperate not only with the present set of companions, those in today’s body. In the next generation, they’ll have to cooperate with a different sample of companions drawn from the shared gene pool. They must be ready to cooperate with all the alternative genes that march with them down the generations within this gene pool – but no other gene pool. This is because Darwinian success, for a gene, means long-term success, travelling through time over many generations, in many successive bodies. They must be good travelling companions of all the genes in the stirred gene pool of the species.
The 1957 film of JB Priestley’s novel The Good Companions had an accompanying song with a not uncatchy tune, of which the refrain was,
Be a good companion, Really good companion, And you’ll have good companions too.
It is a song whose evoked mutualism suits the travelling troupe of genes, which constitutes the active gene pool of a species such as ours. Sexual recombination of genes gives meaning to the very existence of the ‘species’ as an entity worth distinguishing with a name at all. Without it, as is the case with bacteria, there is no distinct ‘species’, no clear way to divide the population with confidence into discrete nameable groups. It is sexual reproduction that confers identity on the species. Some bacterial types are not far from being a big smear, grading into each other as they promiscuously share genes. The attempt to assign discrete species names to such bacteria is a losing battle in a way that doesn’t apply to animals like us, where sexual exchange is limited to sexual encounters between a male and a female of the same species – and no other species by definition. As already stated, where fossils are concerned we have to guess, based on their anatomical similarity, whether they would have been able to interbreed when they were alive. This involves subjective judgement, which is why naming fossils such as Homo rhodesiensis and Homo heidelbergensis is a matter of aggravated controversy between ‘lumpers’ and ‘splitters’. But notwithstanding naming disagreements, which can even become acrimonious, we remain confident that the gene pool surrounding every one of those fossils was a troupe of travelling companions isolated from other gene pools – even though imperfectly isolated during episodes of speciation. Bacteria largely deny us that confidence. So-called ‘species’ of bacteria are not clearly delimited.
Every working gene, ‘expert’ in rendering up its own contribution to the collaborative building of an embryo, is confined to its own gene pool. Repeated cooperation among successive samples drawn from the same troupe of travelling companions has selected genes largely incapable of working beneficially with members of other troupes. Not entirely, as we see from headlined examples like jellyfish genes transplanted into cats and making them glow in the dark. Genes are normally not put to that kind of test. Mules and hinnies, ligers and tigons, are almost always sterile. Their sets of travelling companions are still compatible enough to collaborate in building strong bodies. But their compatibility breaks down when it comes to chromosomal pairing-off in meiosis, the process of cell division that makes gametes. Mules can pull a cart, but they can’t make fertile sperms or eggs.
Nature doesn’t transplant antelope genes into leopards. If it did, a few might work normally. There are broad similarities between the embryologies of all mammals, and all mammals doubtless share genes for making most layers of the mammalian palimpsest. But that doesn’t undermine this chapter. Those genes concerned with what makes a leopard a predator, and an antelope its herbivorous prey, would not work harmoniously together. In childishly crude terms, leopard teeth wouldn’t sit well with antelope guts and antelope feeding habits. Or vice versa. In the language of this chapter, companions that travel well together in one gene pool would not be good companions in the other. The collaboration would fail.
The principle is illustrated by an old experiment of EB Ford, the eccentrically fastidious aesthete from whom I learned my undergraduate genetics. Most practical geneticists work on lab animals or plants, breeding fruit flies or mice in the laboratory. But Ford walked a minority path among geneticists. He and his collaborators monitored evolutionary change in gene pools, in the wild. A lifelong authority on butterflies and moths, he went out into the woods and fields, heaths and marshes of Britain, waving his butterfly net and sampling wild populations. He inspired others to do the same kind of thing with wild fruit flies, wild snails and flowers, as well as other species of butterflies and moths. He founded a whole discipline called Ecological Genetics and wrote the book of that title. The piece of work that I want to talk about here was a field study of wild populations of lesser yellow underwing moths, in Scotland and some of the Scottish islands. Ford knew it as Triphaena comes, but it is now called Noctua comes, following the strict precedence rules of zoological nomenclature.
The species is polymorphic, meaning there are at least two genetically distinct types coexisting in significant proportions in the wild. Not in England, however, nor in much of mainland Scotland, where all the lesser yellow underwings look like the pale upper one in the picture. But in some of the Scottish islands there exists, in significant numbers, a second morph, of darker colour, called curtisii, evidently named after the entomologist and artist John Curtis (1791–1862). I thought it fitting to use Curtis’s own painting of the curtisii morph and the cowslip, and I asked Jana Lenzová to paint in the light morph to complete the picture.
Dark and light morphs of lesser yellow underwing
The difference between the two morphs is controlled by a single gene, which we can call the curtisii gene. Curtisii is nearly dominant. This means that if an individual has either one curtisii gene (‘heterozygous’) or two curtisii genes (‘homozygous for curtisii’), it will be dark. If dominance were complete, heterozygous individuals with one curtisii gene would look exactly the same as homozygotes with two. Curtisii being only nearly dominant, the heterozygotes are almost the same as the curtisii homozygotes but slightly lighter. Heterozygotes are always darker than individuals homozygous for the standard comes gene, which is therefore called recessive.
Like his mentor Ronald Fisher, whom we’ve already met, Ford liked to speak of ‘modifiers’, genes whose effect is to modify the effects of other genes. According to Fisher’s theory of dominance, to which Ford subscribed, when a gene first springs into existence by mutation, it is typically neither dominant nor recessive. Natural selection subsequently drives it towards dominance or recessiveness via the gradual accumulation, through the generations, of modifiers. Dominance is not a property of a gene itself, but a property of its interactions with its companion modifiers.
Modifiers don’t change the major gene itself. What they change is how it expresses itself, in this case its degree of dominance. The language of this chapter would say that a major gene such as curtisii has modifiers among its ‘good companions’, which affect its dominance, meaning its tendency to express itself when heterozygous. For reasons we needn’t go into, natural selection favoured a significant proportion of dark curtisii morphs on certain Scottish islands. And one way this favour showed itself, according to the theory of Fisher and Ford, was by selection in favour of modifiers that increased its dominance.
Barra is an island in the Outer Hebrides, west of Scotland. Orkney, north of Scotland, is an archipelago 340 kilometres from Barra as the crow flies, and too far for the moth to fly. Ford collected and studied moths from both these locations. Both have mixed populations of Lesser Yellow Underwings, the normal pale form living alongside significant numbers of dark curtisii morphs. Breeding experiments, with both Barra and Orkney moths, separately confirmed the dominance of curtisii within both islands. However, when Ford took moths from Barra and crossed them with moths from Orkney, he got a remarkable result. The dominance broke down. It disappeared. No longer did Ford see tidy Mendelian segregation of dark versus light forms. Instead there was a messy spectrum of intermediates. Dominance had disappeared.
What had evidently happened was this. Dominance on Barra had evolved by an accumulation of mutually compatible modifiers, good Barra companions. Dominance on Orkney had independently and convergently evolved by a different consortium of modifier genes, good Orkney companions. When Ford bred across islands, the two sets of modifiers couldn’t work together. It was as though they spoke different languages. To work properly, each modifier needed its normal set of good companions, the set that had been built up over generations of selection on the different islands. That’s what being good companions is all about, and Ford’s experiment dramatically demonstrates a principle that I believe to be general. The ‘major’ gene, curtisii, is the same on both Barra and Orkney. However, for all that a gene itself is the same, its dominance can be built up in more than one way by different consortia of modifiers. This seems to have been the case with curtisii on different islands.
There’s a potential fallacy lurking here. It’s easy to presume that the Barra good companions lie close to each other on a chromosome and therefore segregate as a unit. And likewise, the Orkney consortium of good companions. That kind of thing can happen, and Ford and his colleagues discovered it in other species. Natural selection can favour inversions and translocations of bits of chromosome that bring good companions closer to each other. Sometimes they end up so close that they are called a ‘supergene’, so close that they are rarely separated by crossing over. This is an advantage, and the translocations and inversions that contribute to the building of a supergene are favoured by natural selection. But if Ford’s modifiers had been clustered together as a supergene in the case of his yellow underwings, he wouldn’t have got the results that he did.
Supergenes can be demonstrated in the lab by breeding large numbers of individuals for many generations until suddenly, by a freak of chromosomal crossing-over, the supergene is split. But the supergene phenomenon is not necessary for good companionship, and there’s no reason to suppose it applies in this case of the lesser yellow underwing. The suites of cooperating modifiers could lie on different chromosomes all over the genome. Separately, in their respective island gene pools, they were assembled by natural selection as good team workers in each other’s presence. In this case, they work well together to increase the dominance of the curtisii gene. But the principle is more general than that. We don’t have to subscribe to the Fisher/Ford theory of dominance in particular.
Natural selection favours genes that work together in their own gene pool, the gene pool of their species. Genes that go with being a carnivore (say, genes for carnivorous teeth) are naturally selected in the presence in the same gene pool of other ‘carnivorous genes’ (say genes for short carnivorous intestines whose cells secrete meatdigesting enzymes). At the same time, on the herbivore side, genes for flat, plant-milling teeth flourish in the presence of genes for long, complicated guts that provide havens for plant-digesting micro-organisms. Once again, the alternative suites of genes may be distributed all over the genome. There’s no need to assume that they cluster together on any particular chromosome.
Unfortunately, good companionship sometimes breaks down. It is even subject to sabotage. We’ve already met ways in which the genes within a body can be in conflict with one another. The uneasy pandemonium of genes within the genome, sometimes cooperating, sometimes disputing, is captured in Egbert Leigh’s ‘Parliament of Genes’. Each acts ‘in its own self-interest, but if its acts hurt the others, they will combine together to suppress it.’
Cell division within the body is vulnerable to occasional ‘somatic’ mutation. Of course it is. How could it not be? We are familiar with the idea that random copying errors, mutations, produce the raw material for natural selection between individuals. Those ‘germline’ mutations occur in the formation of sperms and eggs, and they are then inherited by an individual’s children. These are the mutations that play an important role in evolution. But most acts of cell division occur within the body – somatic as opposed to germline mutation – and they too are subject to mutation. Indeed, the mutation rate per mitotic division is higher than for meiotic division. We should be thankful our immune system is so good at spotting the danger early. Most somatic mutations, like most germline mutations, are not beneficial to the organism. Sometimes they are beneficial to themselves but bad for the organism, in which case they may engender malignant tumours – cancers. Subsequent natural selection within the tumour can generate a progression through increasingly ominous ‘stages’ of cancer. I shall return to this.
We can think of the (somatic) cells in a developing embryo as having a family history within the body, springing from their grand ancestor, the single fertilised egg cell of a few months or weeks previous. At any stage in this history of descent, starting with the embryo and on throughout the rest of life, somatic mutation can occur. Vertebrate development is the product of countless cell divisions, so embryologists have found it convenient to trace cell lineages in a simpler organism. The tiny roundworm Caenorhabditis elegans has only 959 cells. It was the genius of the great molecular biologist Sydney Brenner to pick this animal out as the ideal subject for a genre of research that has since spread to dozens of labs throughout the world. Its embryo at one of its developmental stages has precisely 558 cells. Every one of those 558 cells has its own ‘ancestral’ sequence within the developing embryo. The pedigree of each of those 558 cells within the embryo has been painstakingly worked out (illustration below). Necessarily, it’s impossible to print the details legibly on one page of a book, but you can expand it here (https://www.wormatlas.org/celllineages.html) and get an idea of the diverging pedigree of cells in the embryo, consisting of ‘families’ and ‘sub-families’. If you could read the labels by the side of families of cells, you’d see things like ‘intestine’, ‘body muscle’, ‘ring ganglion’. We shall have need to return to that idea of families of cells procreating in the embryo.
Now, if that’s what the cellular pedigree looks like for a mere 558 roundworm cells, just think what it must look like for our 30 to 40 trillion cells. Similar labels – muscle, intestine, nervous system, etc. – could be affixed to cells in a human embryo (opposite). This is true even though the pedigrees are not determined so rigidly in a vertebrate embryo, and we can’t enumerate a finite tally of named cells. It’s important to stress that these different families of cells within the developing embryo are, until something goes wrong, genetically identical. If they weren’t, they might not cooperate. When something goes wrong and they’re no longer genetically identical, well that’s when there’s a risk of their becoming bad companions. And then there’s a risk of their evolving, by natural selection within the body, to become very bad companions indeed: cancers.
As you can see on the diagram on the facing page, after some early cell generations within the embryo, the pedigree of our cells splits into three major families: the ectoderm, the mesoderm, and the endoderm. The ectoderm family of cells is destined to give rise, further down the line, to skin, hair, nails, and those hugely magnified nails that we know as hooves. Ectodermal derivatives also contribute the various parts of the nervous system. The endoderm family of cells branches to give rise to sub-families that eventually make the stomach and intestines; and other sub-families that make the liver, lungs, and glands such as the pancreas. The mesoderm dynasty of cells spawns numerous sub-families, which branch again and again to produce muscle, kidney, bone, heart, fat, and the reproductive organs, although not the germline, which is early hived off and sequestered for its privileged destiny, on down the generations.
Somatic mutants apart, every one of the cells in the expanding pedigree has the same genome, but different genes are switched on in different tissues. That is to say they are epigenetically different while being genetically the same (see the relevant endnote if popular hype has confused you as to the true meaning of ‘epigenetics’). Liver cells have the same genes as muscle cells, but once they pass beyond a certain stage in embryonic development, only liver-specific genes are active there. And the liver ‘family’ of cells in the pedigree goes on dividing until the liver is complete. They then stop dividing. The same applies to all the ‘families’, which each have their own stopping time. Cells must ‘know’ when to stop dividing. And that is where trouble can step in.
With an important reservation, the number of cell generations before the arresting of cell division varies from tissue to tissue and is typically between forty and sixty. That may seem surprisingly few. But remember the power of exponential growth. Fifty liver cell generations, if each one was a division into two (fortunately it isn’t) would yield a liver the size of a large elephant. Different cell lines stop dividing after different limits, producing end organs of different sizes. You can see how important it is for each cell line to know when to stop dividing.
Cactus with somatic mutation
Every one of the 30 trillion cells in a body was made by a cell division. And every one of those cell divisions is vulnerable to somatic mutation. Now we come to that ‘important reservation’, the one relevant to the topic of bad companions. The cells in a lineage are genetically identical only if no somatic mutation intervenes during the lineage’s successive generations. Most somatic mutations are harmless. But what if a somatic mutation arises in a cell such that it changes its behaviour and refuses to stop dividing? Its lineage in the ‘family tree’ doesn’t come to a disciplined halt, but goes on reproducing out of control. The daughter cells of the mutant cell inherit the same rogue mutation, so they too divide. And their daughter cells inherit the rogue gene, so … This is the kind of thing that produces weird growths such as adorns the cactus opposite.
Let’s follow the subsequent history of a rogue cell’s descendants, for example in a human. Reproducing for an indefinite number of generations without discipline, these cells will now be subject to a form of natural selection. Why say ‘a form of’? It is natural selection, plain and simple. The rogue cells will be subject to natural selection, every bit as Darwinian as the natural selection that chooses the fastest pumas or pronghorns, the prettiest peacocks or petunias, the most fecund codfish or dandelions. Rogue somatic mutant cells can evolve, by natural selection within the body, into cancers that spread menacingly (‘metastasis’) to other parts of the body. Now natural selection of cells within the tumour will favour those that become better cancers. What does ‘better’ mean, for a cancer? They become expert, for example, at usurping a large blood supply to nurture themselves. The whole subject, fascinating, disturbing, and not at all surprising to a Darwinist, is expounded in books such as Athena Aktipis’s The Cheating Cell, and The Society of Genes by Itai Yanai and Martin Lercher.
Since cancers evolve by natural selection (within the body), we should treat their evolutionary adaptations in just the same way we might treat the adaptations of pronghorn or codfish, except that the ecological environment is the interior of a (say) human body instead of the sea or an open prairie. This chapter’s discussion of Good Companions has prepared us for the idea of an ecology of genes within the body, to parallel the more conventional idea of an external ecology. And that internal ecology is also the setting where bad companions can thrive. An important difference is that natural evolution in the open sea or prairie goes on into the indefinite future. The evolution of a cancer tumour ends abruptly with the death of the patient, whether that death is caused by the cancer or something else. The cancer evolves to become better and better at (as an inadvertent by-product) killing itself. This, too, should not surprise. Natural selection, as I’ve said over and over, has no foresight. A tumour cannot foresee that increased malignancy will eventually kill the tumour itself. Natural selection is the blind watchmaker. Despite ending with the death of the organism, the number of generations of cell division in a tumour is large enough to accommodate constructive evolutionary change. Constructive from the point of view of the cancer. Destructive for the patient. Athena Aktipis’s book artfully treats the evolution of cancer cells in the body in just the kind of way we might treat the evolution of buffalos or scorpions in the Serengeti.
Cancer cells, then, or rather the mutant genes that turn cells cancerous, are one kind of ‘bad companion’. Another type is the so-called segregation distorter. Sperms and eggs – gametes – are ‘haploid’ cells you’ll remember, having only one copy of each gene, instead of two like normal body cells. The special kind of cell division called meiosis makes haploid gametes (having only one set of chromosomes) out of diploid cells, which have two sets of chromosomes, one set from the individual’s mother and another set from the father. It is only when gametes are made by meiosis that the two sets meet each other in the same chromosome. Meiosis performs an elaborate shuffle, cutting and pasting exchanged portions of paternal and maternal chromosomes into a new set of mixed-up chromosomes. Every gamete is unique, having different assortments of paternal and maternal genes in each of its (twenty-three in humans) chromosomes. The result of the shuffle is that each gene from the diploid set of (forty-six in humans) chromosomes has a 50 per cent chance on average of getting into each gamete.
The ‘phenotypic effect’ of a gene commonly shows itself somewhere in the body – it might affect tail length or brain size or antler sharpness. But what if a gene were to arise that exerted its phenotypic effect on the process of gamete production itself? And what if that effect was a bias in gamete production such that the gene itself had a greater than 50 per cent chance of ending up in each gamete? Such cheating genes exist – ‘segregation distorters’. Instead of the meiotic shuffle resulting in a fair deal to each gamete as it normally does, the deal is biased in favour of the segregation distorter. The distorter gene has a greater than even chance of ending up in a gamete.
You can see that if a rogue segregation distorter were to arise it would tend, other things being equal, to spread rapidly through the population. The process is called meiotic drive. The rogue gene would spread, not because of any advantage to the individual’s survival or reproductive success, not because of benefit of any kind in the conventional sense, but simply because of its ‘unfair’ propensity to get itself into gametes. We could see meiotic drive as a kind of population-level cancer. A special case of a segregation distorter is the ‘driving Y-chromosome’, that is, a gene on a Y-chromosome whose effect on males is to bias them towards producing Y sperms and therefore male offspring. If a driving Y arises in a population, it tends towards driving it extinct for lack of females: population-level cancer indeed. Bill Hamilton even suggested that we could control the yellow fever mosquito by deliberately introducing a driving male into the population. Theoretically, the population should drastically shrink through lack of females.
Other ways have been suggested to control pests by ‘driving genes’. I’ve already mentioned in Chapter 8 the crass irresponsibility of the 11th Duke of Bedford in introducing grey squirrels, native to America, into Britain. He not only released them in his own estate, Woburn Park, but made presents of grey squirrels to other landowners up and down the country. I suppose it seemed like a fun idea at the time, but the consequence is the wiping-out of our native red squirrel population. Researchers are now examining the feasibility of releasing a driving gene into the grey squirrel gene pool. This would not be carried on the Y-chromosome but would produce a dearth of females in a slightly different way. The authors of the idea are mindful of the need to be careful. We want to drive the grey squirrel extinct in Britain but not in America where it belongs and where it would have stayed but for the Duke of Bedford.
Bad companions, at least in the form of cancers, force themselves upon our forebodings. But for our purposes in this book, it is the gene’s role as good companion that we must thrust to prominence. It remains for the last chapter to pin down exactly what makes them cooperate. Fundamentally, it is, I maintain, the fact that they share an exit route from each body into the next generation.
Good companions dressed for field work: RA Fisher and EB Ford. See endnote for my suspicion that this is a historic photogaph.
13 Shared Exit to the Future
Purveyors of scientific wonder like to surprise us with the prodigious – disturbing to some – numbers of bacteria inside our bodies. We’re accustomed to fearing them but most of them are, in the words of Jake Robinson’s title, Invisible Friends. Mostly in the gut, estimates vary from 39 trillion to 100 trillion, the same order of magnitude as the number of our ‘own’ cells, where 40 trillion is a round-number estimate. Between a half and three-quarters of the cells in your body are not your ‘own’. But that doesn’t take account of the mitochondria. These miniature metabolic engine-rooms swarm inside our cells and the cells of all eucaryotes (that is, all living creatures except bacteria and archaea). It is now established beyond doubt that mitochondria originated from free-living bacteria. They reproduce by cell division like bacteria, and each has its own genes in a ring-shaped chromosome, again like bacteria. In fact, let us not mince words, they are bacteria: symbiotic bacteria that have taken up residence in the hospitable interior of animal and plant cells. We even know, from DNA-sequence evidence, which of today’s bacteria are their closest cousins. The number of mitochondria in your body is many trillions.
The bacteria that became mitochondria brought with them much essential biochemical expertise, the research and development of which was presumably accomplished long before they became incorporated as proto mitochondria. Their main role in our cells is the combustion of carbon-based fuel to release needed energy. Not the violent high-speed combustion of fire, of course, but a slow, orderly, trickle-down oxidation. Not only are you a swarm of bacteria, you couldn’t move a muscle, see a sunset, fall in love, whistle a tune, despise a demagogue, score a goal, or invent a clever idea without the unceasing activation of their chemical knowhow, expert tricks cobbled together by natural selection choosing between rival bacteria in a lost pre-Precambrian sea.
The interiors of plant cells swarm with green chloroplasts, which also are descended from bacteria (a different group, the so-called cyanobacteria). Like mitochondria, chloroplasts are bacteria in every sensible meaning of the word. Again like mitochondria, they brought with them a formidable dowry of biochemical wizardry, in this case photosynthesis. Virtually all life on Earth is ultimately powered by energy radiated from the gigantic nuclear fusion reactor that is the sun. It is captured by photosynthesis in chloroplast-equipped solar panels such as leaves, and is subsequently released in the chemical factories that are mitochondria, in all of us. Solar photons that fall on the sea are captured not by leaves but by single-celled green organisms. Whether on land or at sea, solar energy is the base of all food chains. I think the only exceptions are those strange communities whose ultimate source of energy is hot springs, undersea ‘smokers’ and such conduits of heat from the Earth’s interior.
Our mitochondria couldn’t do without us, just as we wouldn’t survive two instants without them. We are joined deep in mutual amity. Our genes and their genes are good companions that have travelled in lockstep over 2 billion years, each naturally selected to survive in an environment furnished by the other. Most of the genes that originated in their bacterial forebears have long since either migrated into our own chromosomes or been laid off as redundant. But why are mitochondria, and some other bacteria, so benign towards us, while other bacteria give us cholera, tetanus, tuberculosis, and the Black Death? My Darwinian answer is as follows. It is an example of the take-home message of the whole chapter. Mitochondrial genes and ‘own’ genes share the same exit route to the future. That is literally true if we are female, or if we for the moment overlook the fact that mitochondria in males have no future. The key to companionable benevolence, I shall show, or its reverse, is the route by which a gene travels from a present body into a body of the next generation.
Mitochondria and chloroplasts may be the earliest examples of bacteria being coopted into animals, but they are not the only ones. Here’s a much more recent re-enactment of those ancient incorporations, and it is highly congenial to the thesis of the gene’s-eye view. The embryonic development of vertebrate eyes requires a protein called IRBP, which facilitates the separation of retinal cells from one another and helps them to see better. In a large survey of more than 900 species, IRBPs were found in every vertebrate examined, plus Amphioxus, a small, primitive creature related to vertebrates, although it lacks a backbone. But of the 685 invertebrate species, the only one with a molecule resembling IRBP was an amphipod crustacean, Hyalella. Among plants, a single species, Ricinus communis, the castor oil plant, has something like an IRBP. And there’s a little cluster of fungi too. Molecules resembling IRBPs are ubiquitous among bacteria.
A family tree of IRBP-like molecules shows a richly branched pedigree among bacteria, paralleling that of the vertebrates in which they live, both pedigrees springing from a single point. The isolated pop-ups (crustacean, fungi, and plant) also spring from within the bacterial tree, but widely separated parts of it. This is good evidence of horizontal gene transfer from various bacteria into the eucaryote genome. The evidence strongly suggests that vertebrate IRBPs are ‘monophyletic’, all descended from a single ancestor, which means a single jump from a bacterium right at the base of vertebrate evolution. Ever since that event, the genes concerned have been passed vertically down the generations. This is like the bacteria that became mitochondria, although mitochondrial ancestors were whole bacteria, not single genes.
I want to give a general name to bacteria that are transmitted from host to host in host gametes: verticobacter, because they pass vertically down the generations. The ancestors of mitochondria and of chloroplasts are prime examples of verticobacters. Verticobacters can infect another organism only by riding inside its gametes into its children. By contrast, a typical ‘horizontobacter’ might pass by any route from host to host. If it lives in the lungs, for instance, we may suppose its method of infection is via droplets coughed or sneezed into the air and breathed in by its next victim. A horizontobacter doesn’t ‘care’ whether its victim reproduces. It only ‘wants’ its victim to cough (or sneeze, or make bodily contact by hands, lips, or genitals), and it works to that end – ‘works’ in the sense that its genes have extended phenotypic effects on the host’s body and behaviour, driving the host to infect another host. A verticobacter, by contrast, ‘cares’ very much that its ‘victim’ shall successfully reproduce, and ‘wants’ it to survive to reproduce. Indeed, ‘victim’ is scarcely the appropriate word, which is why I protected it behind quotation marks. This is, of course, because a verticobacter’s ‘hope’ of future transmission lies in the offspring of the host, exactly coinciding with the ‘hopes’ of the host itself. Therefore, if a verticobacter’s genes have extended phenotypic effects on the host, they will tend to agree with the phenotypic effects of the host’s own genes. In theory a verticobacter’s genes should ‘want’ exactly the same thing as the host’s genes in every detail.
The pertussis (whooping cough) bacterium is a good example of a horizontobacter. It makes its victims cough, and it passes through the air to its next victim, in droplets emitted by the cough. Cholera is another horizontobacter. It exits the body via diarrhoea into the water supply, whence it ‘hopes’ to be imbibed by somebody else, drinking contaminated water. It doesn’t ‘care’ if its victims die, and it has no ‘interest’ in their reproductive success.
The notion of a parasite’s ‘wanting’ its victim to do something needs explaining, and this again is where the extended phenotype comes in, as promised at the end of Chapter 8. The parasitology literature is filled with macabre stories of parasites manipulating host behaviour, usually changing the behaviour of an intermediate host to enable transmission to the next stage in the parasite’s complicated life cycle. Many of these stories concern worms rather than bacteria, but they convey the principle I am seeking to get across. ‘Horsehair worms’ or ‘gordian worms’, belonging to the phylum Nematomorpha, live in water when adult, but the larvae are parasitic, usually on insects. The insect hosts being terrestrial, the gordian larva needs somehow to get into water so it can complete its life cycle as an adult worm. Infected crickets are induced to jump, suicidally, into water. An infected bee will dive into a pond. Immediately the gordian worm bursts out and swims away, the crippled bee being left to die. This is presumably a real Darwinian adaptation on the part of the worm, which means that there has been natural selection of worm genes whose (‘extended’) phenotypic effect is a change in insect behaviour.
Here’s another example, this time involving a protozoan parasite, Toxoplasma gondii. The definitive host is a cat, and the intermediate host is a rodent such as a rat. The rat is infected via cat faeces. Toxoplasma then needs the infected rat to be eaten by a cat, to complete its life cycle. It insinuates itself into the rat’s brain and manipulates the rat’s behaviour in various ways to that end. Infected rats lose their fear of cats, specifically their aversion to the smell of cat urine. Indeed, they become positively attracted to cats, though apparently not to non-predatory animals, or predators that don’t attack rats. There is some evidence that they lose fear in general, owing to increased production of the hormone testosterone. Whatever the details, it’s reasonable to guess that the change in rat behaviour is a Darwinian adaptation on the part of the parasite. And therefore an extended phenotype of Toxoplasma genes. Natural selection favoured Toxoplasma genes whose extended phenotypic effect was a change in rat behaviour.
The infected snail’s bulging eyes are a tempting target for birds
Leucochloridium is a fluke (flatworm), parasitic on birds. Its intermediate host is a snail, and it needs to transfer itself from snail to bird. The snails that it infects are largely nocturnal, while the birds who are the next host feed by day. The worm manipulates the behaviour of the snail to make it go out by day. But that is only the beginning of the snail’s troubles. One of the life-history stages of the worm invades the eye stalk of the snail, which swells grotesquely, and seems to pulsate vividly along its length.
This is said to make the eye stalk look like a little crawling caterpillar. Be that as it may, it certainly renders the eye stalks conspicuous, and birds readily peck them off. Infected snails also move around more actively than unparasitised ones. The snail is not killed but only blinded. It is able to regenerate its eye stalks to pulsate another day and perhaps be again plucked off. The fluke, for good measure, castrates its snail victim. And that’s an interesting story in its own right. ‘Parasitic castration’ is common enough to be a named thing. It is practised by a wide variety of parasites from around the animal kingdom, including protozoa, flatworms, insects, and various other crustaceans. Including Sacculina, the parasitic barnacle that I introduced in Chapter 6 and promised to return to.
Sacculina is perhaps the most extreme example of the ‘degenerative’ evolution typical of parasites. Darwin, in his monographs on barnacles, which distracted him for eight of the twenty years when he might have published on evolution, misdiagnosed the affinities of Sacculina. And who can blame him? Just take a look at it. The externally visible part of Sacculina is a soft bag clinging to the underside of a crab. Most of the ‘barnacle’ consists of a branching root system permeating the inside of the unfortunate crab’s body. Eventually, it fills the body so completely that if you could sweep away the crab and leave only the Sacculina, this is what you might see.
This is not a crab
How do we know that this system of branching rootlets, this sprawling entity that looks like a plant or fungus, is really a barnacle? How do we even know it’s a crustacean? The various larval stages of the life cycle give it away. The nauplius larva is followed by the cyprid larva, and both are unmistakeably crustacean. As if final clinching were needed, Sacculina’s genome has now been sequenced. ‘It is written’, as the Muslims say: ‘Crustacean’.
Sacculina larvae
The first organs that Sacculina attacks are the crab’s reproductive organs. This is the ‘parasitic castration’ that I mentioned above. Barnacles themselves are sometimes castrated by parasitic crustaceans; marine isopods related to woodlice. So, what is the point of parasitic castration? Why would a parasite head straight for the gonads of its host, before eating other organs?
As with all animals, the host’s ancestors have been naturally selected to fine-tune a delicate balance between the need to reproduce (now) and the need to survive (to reproduce later). A parasite such as Sacculina, however, has no interest in assisting its host to reproduce. This is because its genes don’t share the host genes’ exit route to the future. Sacculina genes ‘want’ to shift the host’s ‘balance’ towards surviving, to carry on feeding the parasite. Like a docile, castrated ox being fattened up, the crab is forced by the parasite to renounce reproduction and become a maintained source of food.
The situation reverses in those cases where parasites – ‘verticoparasites’ – pass to the next host generation in the gametes of the host. Verticoparasites infect only the offspring of their individual hosts rather than potential hosts at large. The genes of a verticoparasite share the ‘exit route’ of the host genes, so their extended phenotypic effects will agree with the host genes’ phenotypic effects. Exercise our usual cautious licence to personify, and consider the ‘preferred options’ of a verticoparasite such as a verticobacter. It travels inside the eggs of a host directly into the host’s child. Here, the interests of parasite and host coincide, and their genes ‘agree’ about the optimal host anatomy and behaviour. Both ‘want’ the host to reproduce, and survive to reproduce. Once again, if the genes of vertically transmitted parasites have extended phenotypic effects on their hosts, those effects should coincide, in perfect agreement and in every detail, with the phenotypic effects of the host animal’s ‘own’ genes.
Mitochondria are an extreme example of a verticoparasite. Long transmitted vertically down the generations inside host eggs, they became so amicably cooperative that their parasitic origins are hard to spot, and were long overlooked. A horizontoparasite such as Sacculina has opposite ‘preferences’. It has no ‘interest’ in its host’s successful reproduction. Whether or not a horizontoparasite ‘cares’ about its host’s survival depends on whether it can benefit from it, presumably, as in the case of Sacculina, by feeding on the living host. If, by castration, it can shift the balance of the host’s internal economy away from reproduction and towards survival, so much the better.
The tapeworm Spirometra mansanoides doesn’t castrate its mouse victims but it achieves a similar result. It secretes a growth hormone, which makes them grow fatter than normal mice. And fatter than the optimum achieved by natural selection of mouse genes seeking a balance between growth and reproduction. Tribolium beetles normally develop through a succession of six larval moults, increasing in size, before they eventually change into an adult. A protozoan parasite, Nosema whitei, when it infects Tribolium larvae, suppresses the change to adult. Instead, the larva continues to grow through as many as six extra larval moults, ending up as a giant grub, weighing more than twice as much as the maximum weight of an uninfected larva. Natural selection has favoured Nosema genes whose extended phenotypic effect was a dramatic doubling in Tribolium fatstock weight, achieved at the expense of beetle reproduction.
A small tapeworm, Anomotaenia brevis, needs to get into its definitive host, a woodpecker. It does so via an intermediate host, an ant of the species Temnothorax nylanderi, which has the habit of collecting woodpecker droppings to feed to its larvae. Tapeworm eggs are often present in the droppings, and can therefore find themselves being eaten by ant larvae. The parasite then has an interesting effect on the ant’s behaviour when it becomes adult. It refrains from work and is fed by unparasitised workers. Parasitised ants also live longer, up to three times longer, than normal ants. This increases their chance of being eaten by a woodpecker – which benefits the tapeworm.
There are parasitic flukes who persuade their snail victim to develop a thicker shell than normal. Shells are presumably an adaptation to protect the snail and prolong its life. But a shell, like any other part of the body, is costly to make. In the personal economics of snail development, the price of thickening the shell is presumably paid out of non-shell pockets, such as those committed to reproduction. Natural selection of snails has built up a delicate balance between survival and reproduction. Too thin a shell jeopardises survival. Too thick a shell, although good for survival, takes economic resources away from reproduction. The fluke, not being a vertically transmitted parasite, ‘cares nothing’ for snail reproduction. It ‘wants’ the snail to shift its priorities towards individual survival. Hence, I suggest, the thickened shell. In extended phenotype language, natural selection favours genes in the fluke that exert a phenotypic effect on the snail, upsetting its carefully poised balance. The thickening of the shell is an extended phenotype of fluke genes, benefiting them but not the snail’s own genes. This case is interesting as an example of a parasite apparently – but only apparently – doing its host a good turn. It strengthens the snail’s armour and perhaps prolongs its life. But if that were really good for the snail, the snail would do it anyway, without the ‘help’ of a parasite. The snail balances a finely judged internal economy. Too lavish spending on survival impoverishes reproduction. The parasite unbalances the snail’s economy, pushing it too far in the direction of survival at the expense of reproduction.
According to the gene’s-eye view of life that I advocate, genes take whatever steps are necessary to propagate themselves into the distant future. In the case of ‘own’ vertically transmitted genes, the steps taken are phenotypic effects on the form, workings, and behaviour of ‘own’ bodies. Genes take those steps because they inherit the qualities of an unbroken, vertically travelling line of successful genes that took the same steps through the ancestral past – that is precisely why they still exist in the present. All of our ‘own’ genes are good companions that agree with each other about what the best steps are. Everything that helps one member of the genetic cartel into the next generation automatically helps all the others. All ‘agree’ about the goal of whatever it is they variously do to affect the phenotype. And why do they agree? Precisely because, in every generation, they share with each other the same exit route into the next generation. That exit route is the gametes – the sperms and eggs – of the present generation. And now we return to verticobacters and other verticoparasites. They have exactly the same exit route as the host’s own genes, and therefore exactly the same interests at heart.
The genes of a verticobacter look back at the same history of ancestral bodies as its host’s own genes. Verticobacter genes have the same reason to behave as good companions towards our own genes as our own genes have towards each other. If an animal benefits from fast-running legs and efficient lungs for running, then its internal verticobacters will also benefit from the same things. If a verticobacter has an extended phenotypic effect on running speed, that effect will be favoured only if it is positive from the organism’s point of view too. The interests of host and bacterium coincide in every particular. A horizontobacter, on the other hand, might be more likely to ‘want’ its victim, when pursued, to cough with exhaustion – coughing being exactly what the horizontobacter needs in order to get itself passed on to another victim. Or another horizontobacter might want its victim to mate more promiscuously than the optimum ‘desired’ by the host’s own genes, thereby maximising contact with another host, and hence opportunities for infection. An extreme horizontobacter might devour the host’s tissues completely, reducing it to a bag of spores which eventually bursts, scattering them to the winds, where they may find fresh hosts to conquer.
A verticobacter ‘wants’ its victims to reproduce successfully (which means, as we saw earlier, that ‘victims’ is not really an appropriate word). Its ‘hopes’ for the future precisely coincide with those of its host. Its genes cooperate with those of the host to build a strong body surviving to reproductive age. Its genes help to endow the host with whatever it takes to survive and reproduce; with skill in building a nest, diligence in gathering food for the infants, success in fledging them at the right time to prepare to reproduce the next generation, and so on. If a verticobacter happens to have an extended phenotypic effect on a host bird’s plumage, natural selection could favour verticobacter genes that brighten the feathers to make the host more attractive to the opposite sex. Verticobacter genes and host genes will ‘agree’ in every respect.
Exactly the same argument applies to viruses, of course. And now we approach the twist in the tail of this chapter and this book. Any virus that travels from human (for example) generation to generation via our sperms or eggs will have the same ‘interests’ as our ‘own’ genes. Whatever colour, shape, behaviour, biochemistry is best for our ‘own’ genes will also be best for (let’s call them) verticoviruses. Verticovirus genes will become good companions of our own genes, accounting for the familiar fact that viruses can help us as well as harm us. Horizontovirus genes, by contrast, don’t care if they kill their victims, so long as they get passed on to new victims by their route of choice – coughing, sneezing, handshaking, kissing, sexual intercourse, whatever it is.
A good example of a horizontovirus is the rabies virus. It is transmitted via the foaming saliva of its victims, whom it induces to bite other animals thereby infecting their blood. It also leads its victims, for example ‘mad’ dogs, to roam far and wide (and out in the midday sun), rather than stay, perhaps sleeping, within their normal home range. This helps the virus by spreading it over a larger geographical area.
What would be a good real example of a verticovirus? It has been estimated that about 8 per cent of the human genome actually consists of viral genes that have, over the millions of years, become incorporated. Among these ‘retroviruses’, some are inert but others have effects that are beneficial. For example, it has been suggested that the evolutionary origin of the mammalian placenta was the result of a beneficial cooperation with an ‘endogenous’ retrovirus that succeeded in writing itself into the nuclear DNA. LP Villarreal, a leading virologist, has gone so far as to suggest that ‘viruses were involved in most all major transitions of host biology in evolution’, and ‘From the origin of life to the evolution of humans, viruses seem to have been involved … So powerful and ancient are viruses, that I would summarize their role in life as “Ex virus omnia” (from virus everything).’
And now, can you see where I am finally going in this chapter? In what sense are our ‘own’ genes different from benign, good companion viruses? Why not push to the ultimate reductio? Why not see the entire genome as a huge colony of symbiotic verticoviruses? This is not a factual contribution to the science of virology. Nothing so ambitious. It’s more like an expansion of what we might mean by ‘virus’ – rather as ‘extended phenotype’ was an expansion of what we might mean by ‘phenotype’. Our ‘own’ genes are verticoviruses, good companions held together and cooperating because they share the same exit route to the next generation. They cooperate in the shared enterprise of building a body whose purpose is to pass them on. Viruses as we normally understand the word, and computer viruses, are algorithms that say ‘Duplicate me’. An elephant’s ‘own’ genes are algorithms that say, in the words of an earlier book of mine, ‘Duplicate me by the roundabout route of building an elephant first’. They are algorithms that work only in the presence of the other genes in the gene pool. They are equivalent to an immense society of cooperating viruses.
I’m not just saying that our genome consists of ‘endogenous retroviruses’ (ERVs) that were once free, infected us, and then became incorporated into the chromosomes. That is true in some cases and it is important, but it’s not what this final chapter is suggesting. Lewis Thomas also didn’t mean what I now mean, although I would love to borrow his poetic vision in pushing the climax of my book.
We live in a dancing matrix of viruses; they dart, rather like bees, from organism to organism, from plant to insect to mammal to me and back again, and into the sea, tugging along pieces of this genome, strings of genes from that, transplanting grafts of DNA, passing around heredity as though at a great party.
The phenomenon of ‘jumping genes’, too, is congenial to my vision of a genome as a cooperative of verticoviruses. Barbara McClintock won a Nobel Prize for her discovery of these ‘mobile genetic elements’. Genes don’t always hold their place on a particular chromosome. They can detach themselves, then splice themselves in at a distant place in the genome. Some 44 per cent of the human genome consists of such jumping genes or ‘transposons’. McClintock’s discovery of jumping genes conjures a vision of the genome as a society, like an ants’ nest: a society of viruses held together only by their shared exit route, and hence shared future and shared actions calculated to secure it.
My suggestion is that the important distinction we need to make is not ‘own’ versus ‘alien’ but vertico versus horizonto. What we normally call viruses – HIV, coronaviruses, influenza, measles, smallpox, chickenpox, Rubella, rabies – are all horizonto viruses. That, precisely, is why many of them have evolved in a direction that damages us. They pass from body to body, via routes that are all their own, by touch, in the breath, by genital contact, in saliva, or whatever it is, and not via the gametic routes with which our own genes traverse the generations. Viruses that share the same genetic destiny as our own genes have no reason to dissent from good companionship. On the contrary. They stand to gain from the survival and successful reproduction of every shared body they inhabit, in exactly the same way as our own genes do. They deserve to be considered ‘our own’ in an even more intimate sense than mitochondria, for mitochondria pass down the female line only. And, from this point of view, our ‘own’ genes are no more ‘own’ than a retrovirus that has become incorporated into one of our chromosomes and stands to be passed on to the next generation by exactly the same sperm or egg route as any other genes in the chromosome.
I cannot emphasise strongly enough that I am not suggesting that all our genes were once independent viruses that later ‘came in from the cold’ and, as retroviruses, ‘joined the club’ of our own nuclear genome. That is known of some 8 percent of our genes, it may be true of many more, it is interesting and important, but it is not what I am talking about here. My point is rather to downplay the distinction between ‘own’ and ‘other’, and to emphasise instead the distinction between vertico and horizonto.
Our entire genome – more, the entire gene pool of any species of animal – is a swarming colony of symbiotic verticoviruses. Once again, I’m not talking only about the 8 percent of our genome that consists of actual retroviruses, but the other 92 percent as well. They are good companions precisely because they are vertically transmitted, and have been for countless generations. This is the radical conclusion towards which this chapter has been directed. The gene pool of a species, including our own, is a gigantic colony of viruses, each hell-bent on travelling to the future. They cooperate with one another in the enterprise of building bodies because successive, temporary, reproduce-and-then-die bodies have proved to be the best vehicles in which to undertake their vertical Great Trek through time. You are the incarnation of a great, seething, scrambling, time-travelling cooperative of viruses.
另一个有趣的化石来自地猿(Ardipithecus),其年代更近,保存也更完整。美国古人类学家蒂姆·D. 怀特(Tim D. White)对埃塞俄比亚多个发掘点出土的数以千计的整骨和碎骨进行了长达15年的精心研究,随后于2009年对这些可追溯到440万年前的地猿化石进行了解读。根据地猿化石周围的动物化石推断,地猿生活在森林里,身高约1.2米,既能行走又能攀缘。地猿长有对生的大脚趾,但不如黑猩猩的灵活。虽然双腿移动起来比大猩猩还要容易一些,但是地猿的双臂和指骨长而弯曲的手指非常适于树栖生活。地猿的犬齿强健有力,具有明显的祖先特征(直接遗传自祖先),脑容量接近黑猩猩。一些人认为地猿是南方古猿(和人类)的直系先祖,另一些人则将地猿视为远房表亲,与黑猩猩的亲缘关系更近。
在布里吉特·森努特和苏珊娜·K. S. 索普(SusannahK. S. Thorpe)等众多古人类学家看来,树栖(指一生中的大部分时间都栖息在树上)的人科动物或许最先发展出了双足行走的特征。我们的直系祖先恰恰生活在森林里,它们应该不是四足行进的,且极有可能习惯于攀缘!我们已经发现,作为现存树栖特征最为明显的猿类,猩猩在踏上柔软树枝时会尽量增加腿部的伸展幅度,与人类在有弹性的地面上奔跑时的肢体反应别无二致,而其他猴子的做法却恰恰相反。由此推断,地面上的双足行走应该是由树上的双足行走发展出来的。古生物学家称此现象为在树上进行的“直立姿势预适应”。
这幅从猴到人的行进图来自时代生活图书公司(Time-Life Books)出版的图书《早期人类》(The Early Man)。毋庸置疑,这幅插图在普及演化思想方面确实发挥了作用。可不幸的是,它在多个层面上都传达了错误的信息。首先,这幅插图给人的感觉是,这些灵长动物无一例外地朝着智人的方向前进,仿佛成为人类是它们不可避免的终极演化结果。其次,插图里的几个物种,并非一个就是另一个的后代:人类的演化不是直进式的,而是分支式的,在演化的过程中,许多物种都消失在了历史长河里,并没有留下任何后代。
1997年,古人类学家罗纳德·J. 克拉克(Ronald J. Clark)在斯泰克方丹发现了一具近乎完整的南方古猿非洲种(或邻近物种)的骨架,并将其命名为“小脚”,其生活年代距今370万年。罗纳德·J. 克拉克认为,“小脚”是雌性古猿,身高约1.3米,去世时约30岁。由于骨骸被封存在极其坚硬的矸石之中,人们在20年之后才将它取出,直至2017年才对它进行了描述。
在“象坑”(Sima del Elefante)里,出土了几个人亚族骸骨化石和一些人亚族活动的痕迹,还有几件奥杜韦风格的工具。经测定,这些骸骨化石的年代为距今120万年,被认为是直立人。与此相比,邻近的格兰多利纳(Gran Dolina)洞穴的骸骨更加丰富,共出土了距今78万年的约80件骨骼碎片。由于与直立人的骨骼稍有不同,这些骨骼碎片被命名为前人(Homo ante-cessor,又称先驱人)。前人使用的工具也是砾石砍砸器,没有一丁点儿两面器的特点。2013年,在英国黑斯堡(Happisburgh)的海滩上发现了同一年代的脚印,研究认为是两个成年原始人和几个儿童留下的。
凭着另一个年代更近的矿层,阿塔普埃尔卡挖掘点享誉世界,这便是被称为Sima de los Huesos的“骨坑”。自1984年开始挖掘以来,坑中已经出土了几个完整颅骨和6800余件其他骨骼,它们属于距今大约43万年的至少28个个体。今天,这些古人类被命名为海德堡人(Homo heidelbergensis),其身材高大强壮,男性平均身高为1.7米,女性平均身高为1.57米,男女之间的身高差与我们差不多。海德堡人能够制造阿舍利风格的两面器。
另一些在欧洲发现的年代更近的化石也表现出类似的特征,比如亨利·德·拉姆利(Henry de Lumley)团队于1969年在法国东比利牛斯省阿拉戈(Arago)洞穴发现的距今45万年至30万年的陶塔维尔人(TautavelMan),或在德国发现的施泰因海姆(Steinheim)颅骨和在希腊发现的佩特拉罗纳颅骨。许多专家都将它们视为海德堡人。在一些研究人员看来,海德堡人是严格意义上的欧洲人种,是从至少100万年前来到欧洲的直立人发展而来的“前尼安德特人”。在演化过程中,这些直立人想必经过了前人阶段。
进入智人体内的新基因随后发生了突变并改变了序列,进而变得与尼安德特人的初始基因有所不同。通过研究突变的数量,就能够确定杂交发生的年代。研究结果表明,杂交可能发生在大约10万年前,那时两者在近东地区比邻而居;抑或是在大约6万年前至5万年前,原先留在非洲的智人最终走出非洲之时。因此,根据4.5万年前生活于西伯利亚的一个智人的DNA研究结果,他的先祖曾在他出生前1.3万年至0.7万年就已经经历过杂交。罗马尼亚的欧亚瑟洞(Pestera cu Oase,意为“骨头洞”)里出土了生活于约4万年前的智人骸骨(这是欧洲已知最古老的智人),他体内的尼安德特DNA占比是现代人的3倍;从他往前追溯就能发现,他4至6代前的祖先还是尼安德特人!不过,这个智人似乎没有留下后代,因为现代人的基因组里已经没有了他的遗传特征。