第六章 与百万年历史决裂
耶路撒冷东北25千米的Tell el Sultan是《圣经》上反复出现的一个地名,1870年代,考古发掘证实,这里实际上属于《圣经》中提及次数更多的古城耶利哥(Jericho)的一部分。后来,这一带陆续出土的古人类定居的遗址逐渐超过20处,时间大多超过4 000年。1952-1958年,英国的女考古学家凯瑟琳·凯尼恩(Kathleen Kenyon,1906-1978)主持对新的土层进行系统挖掘,彻底改变了人们对历史的看法:耶利哥的早期人类遗迹超过一万年。
凯瑟琳·凯尼恩发现:不同于石器时代的先祖,耶利哥(大约7 000年前,人类放弃了这个定居将近4 000年的城市)的人类社区已经开始出现早期农业种植的遗迹,在颅骨上涂抹石膏装饰说明在这里出现了古代崇拜的宗教信仰。现代技术测定确认,这个地区的历史超过9 500年。
1968年,人们在叙利亚境内的幼发拉底河上建造塔巴水坝(Tabqa Dam)时,挖掘出一个人类居住了近4 000年(1.1万——0.75万年前)的遗址——阿布·胡列伊拉(Tell Abu Hureyra)。这是一个从狩猎采集生活形态向农业种植形态过渡的遗址,这里的生活者也因此被称为世界上最早的农民。在这个遗址,从土壤和动物鱼骨等物质中成功分离出712个种子样本,最终查明属于150类以上食用植物的500多种植物种子。这个叙利亚遗址再现了1.1万年前的人类采集狩猎生活方式,和大约一万年前开始的初步的农业种植生活的轮廓。
学者将这种半定居生活的中东文明命名为纳图夫文明(Natufian culture),时间在1.25万——0.9万年,生活在这一带的人群被称为纳图夫人(Natufian)。这个名称的来源,也是出自一位英国女考古学家。1924年,英国女考古学家多萝西·加罗德(Dorothy Garrod)在以色列的纳图夫河道(Wadi an-Natuf)的洞穴里第一次发现了这种文明。
在大约1.25万年前冰河期结束时的温暖的地中海沿岸,仅仅过了几百代,纳图夫人(Natufian)就开始酝酿着一场与上百万年的狩猎采集生活的决裂。人类迁移到中东地区后发现,即使定居在一个地点,也可以采集到足够的植物种子生存下去,于是开始了半定居或定居的纳图夫文明。
大约1.1万年前,气候更加温暖干燥,植物种子产量开始减少。纳图夫人的生活出现了双重的压力:获得更多种子的压力和不愿意脱离舒服的定居生活的压力。但是人们很快想出了解决办法。在不到1 000年的时间里,中东地区的各个定居点逐渐转向植物种子的种植,并且开始利用水利进行灌溉——农业出现了。戈登·柴尔德(Gordon Childe,1892-1957,考古学家和语言学家)把这场巨变命名为“新石器革命”。
农业文明的出现
考古学可以给出细节,解答很多疑问。在追寻人类旅程的过程中,考古学、人类学、语言学、遗传学、生物学组成了同盟军,必须互相借鉴对方的成果。基因技术的出现并未否定其他学科,基因把零零散散的引起多年争议的少量化石证据连接成为一个清晰的证据链。
旧石器时代(Paleolithic)和新石器时代(Neolithic)都被称为石器时代(Stone Age),这两个名称都有后缀lithic(希腊语lithos的意思是石头)。警察经常在垃圾箱里翻腾寻找犯罪证据,考古学家的工作,其实与翻腾垃圾箱差不多。他们翻腾出来在250万——300万年里,人类和其他人属生物制作的无数石器证据,形成了考古学的专业学科和很多学派。
在过去的250万——300万年里,人类的全部技术的99%体现在石器上,所以考古学家能够准确推测出很多历史细节。与石器时代的所有其他的类人生物不同,只有我们人类这个物种,进入了一种新的生活方式,即农业时代。
新石器时代(Neolithic)是人类历史的转折点。从此,人类不再受气候控制,反过来开始控制自己的命运。
第一,农业使人类有了选择的权利,耶利哥的纳图夫人不必每天走十几到几十千米去采集植物的种子。早期农业在中东、中国和美洲分别发生了,人类开始直接控制食物来源。
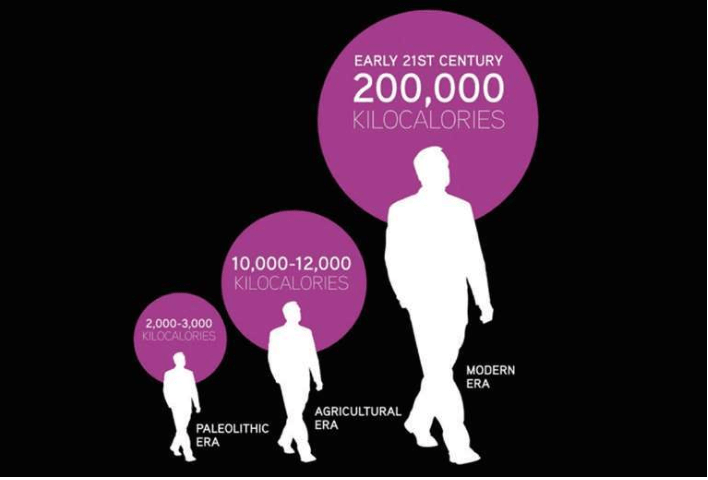
第二,人口开始迅速增长。人类学家虽然并不知道农业出现之前的旧石器时代的人口总数,但估算出农业出现时全球人口仅约几百万,而到了1750年工业革命时,全球人口已约5亿。
第三,农业是人类迁移过程中出现的最新技术。人类曾经在巨大的欧亚干草原带利用新的狩猎技术生存了两万年,而后一个一万年是人类在全球站稳脚跟的时代。
狩猎技术,曾经引发第一次人口迁移,人类开始全球分布。
农业技术,即将引发第二次人口迁移。
20世纪,研究者在中东地区进行了大量的考古挖掘,证实当时这里发生了一场新石器时代革命,时间约为一万年前。新石器革命的发生非常突然,而且是在多处同时发生。
这场革命的起源在土耳其东部的卡拉卡山(Karacadag),科学考察确认至今还有68种野生植物继续生长在这个山区,而且,现在全球食用量最高的小麦最有可能是在卡拉卡山区被驯化出来的。
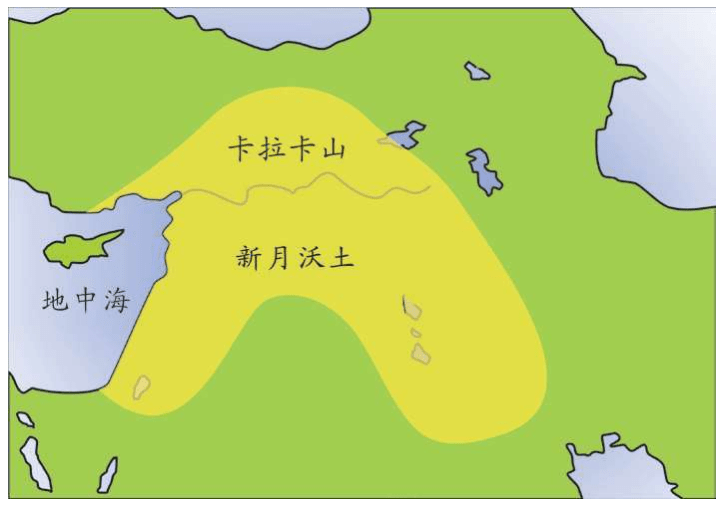
卡拉卡山区丰富的可食用植物种子和种植技术,沿着黎巴嫩——以色列——叙利亚——伊拉克,一直传播到地中海沿岸。其中最著名的遗迹包括耶利哥(Jericho)、叙利亚的阿布·胡列伊拉遗址(Tell Abu Hureyra)、土耳其的加泰土丘(Catal Huyuk)。(加泰土丘挖掘时间在1950-1990年,新石器时代的14层遗迹厚度达15米,时间为8 850年前,当时已经进入所谓的金石混用时代(中东学者对青铜时代的称呼),艺术和宗教的文物极其丰富)
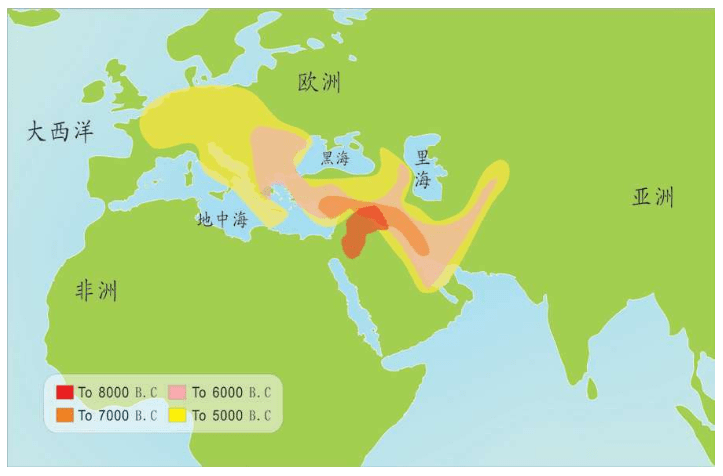
中东地区开始的农业,在亚洲也出现了。在欧洲,南部的农业起源于7 000年前,北部起源于5 000年前。
起源于中东新月沃土(F ertileCrescent)的农业,几千年后传到欧洲。但是这一理论始终存在争议。到底是农业技术传播到欧洲取代了狩猎采集的生活形态,还是中东人群带着农业技术取代了当地的欧洲人?这是完全不同的两个概念。1970年代,斯福扎和另外两个遗传学家Alberto Piazza、Paolo Menozzi开始研究农业对遗传的效应——农业是怎么传播的?
显然,农业在当时是一种“时髦文化”。
这项研究没有取得什么结果。血型和细胞表面蛋白质标记无法确认人的血统世系,也无法落实迁移路线。这是当时的研究技术的限制。但是,斯福扎发现农业并非单纯的文化现象,而是伴随着人口的快速增长,这股风潮从欧洲的东南部向西北部扩散,后来被称为“前进的浪潮”(Wave of Advance)。这种“前进的浪潮”被很多人接受了,但是斯福扎本人并不接受这种观念,因为人们还没有搞清楚欧洲的基因库的起源。
1990年,牛津大学的马丁·理查兹(Martin Richards)和他的同事,开始发表欧洲和东南亚人群线粒体DNA的一系列研究成果。他们最先提出一种小心翼翼的求证绝对时间的方法,再按照绝对时间,估算出欧洲基因库的各种血统迁移的相对分布形态。他们的研究结果得出一种猜测:没有很多农业人口从中东迁移到欧洲,因为看不到这种基因效应,欧洲大部分人口的血统已经在欧洲存在2万——4万年。
2000年,欧尔奈拉·塞米诺(Ornella Semino)等人检测分析了1 000多个欧洲人和中东人的Y染色体,希望找出农业扩张的原因。他们发现,Y染色体的结果与线粒体DNA的研究结果一样,现代欧洲的血统里,中东的基因标记很少——欧洲原来的M173占80%,来自中东的M172仅占20%。但是并非农业没有产生影响,农业传播带来沿途人口的激增。此外,他们计算出3万——1.5万年,欧洲人口曾大幅度下降,当时欧洲正在逐渐步入冰河期最严酷的时代。大约1.6万年前,欧洲人后撤并局限在伊比利亚半岛——意大利南部——巴尔干半岛地区。冰河期结束后,少量幸存者的人数开始上升。
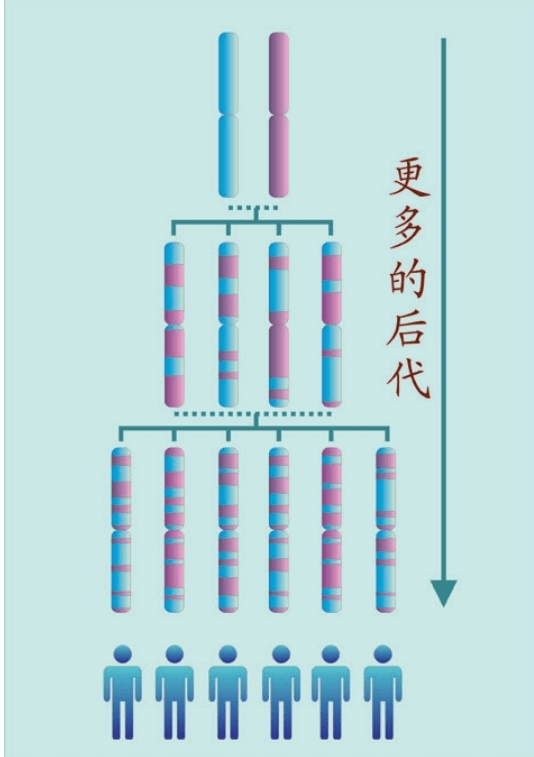
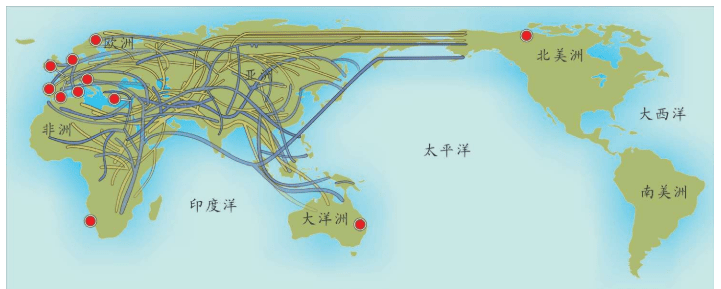
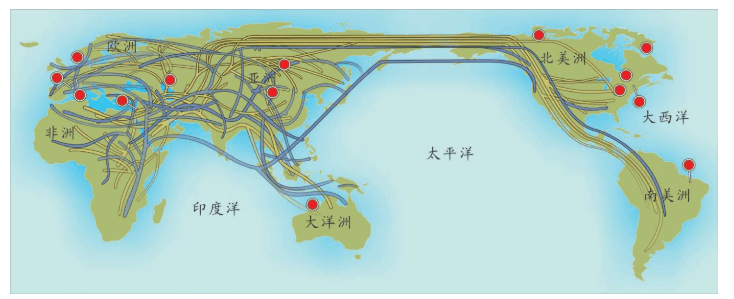
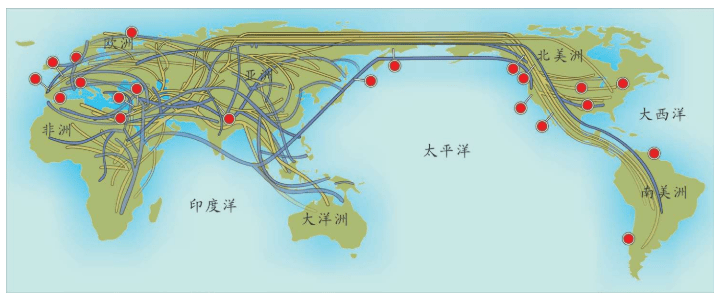
也就是说,基因分析证明:欧洲的农民起源分为两种,大部分农业人口是欧洲原住民,他们学习和接受了农业。欧洲人,尤其欧洲北部的人群自己学来了时髦的农业文明。中东人,喜欢故乡的温暖,没有大规模移民,只有少数中东农民带着农业技术移民来到南欧,这一部分新来者带来了中东的基因,如下图所示。
大量化石记录证明,人类和其他人科生物一样,长期依赖狩猎和采集为生。追随着猎物群体迁移或季节变换,人类也从一个地方游荡到另一个地方。转换为农业和定居生活方式之后,人类的健康状态和社会状态并非是完全正面的,很多证据已经证明,人类的健康和社会反而更槽糕。但是,农业保证了食物供应,带来的一个最大好处是生产人口。
大约一万年前,世界各地的所有狩猎采集人口合计才只有几百万,相当于现在的一座大城市的人口。原本分散的耕地,很快连接成大片耕地,人口得以持续增长。现在,世界人口已达70亿人,增长了1 000倍。
这就是农业革命的结果。
双刃的镰刀
中东人驯化植物的过程比较曲折,亚洲人驯化植物的过程却相对清晰。
中国的植物品种没有中东那么多(新月沃土驯化的农作物为31种,印度——中国——东南亚合计11种),9 000年前黄河流域的主要农作物是粟米,扩散到中国北方其他区域。长江流域的湖南彭头山遗址(Pengtoushan)显示,大约早了两千年,水稻在中国南方被独立驯化出来,然后向长江流域扩散,大约7 000年前成为中国南部的主要农作物,5 500年前传播到中国台湾,4 000年前传播到婆罗洲和苏门答腊,3 500年前传播到印度尼西亚全境。
与欧洲当地人自己学会了农业不同,亚洲地区发生了农业人口的大迁移。在亚洲,水稻技术与基因标记同时传播。如果说,欧洲农业技术带来了“前进的浪潮”(Wave of Advance),那么携带水稻技术的东亚人的大迁移就像一场海啸。
M122是定义东亚氏族的M175上出现的第一个基因标记,在亚洲超过一半的男性Y染色体上都发现了M122。3万年前,M122首次出现。在中亚没有发现M122,在中东和欧洲从来没有发现……但是在东亚,在中国南部、中国北部、日本、太平洋塔希提群岛都发现了M122,而且扩散速度非常快。
M122,现在定义了中国农业人的后裔。
大卫·戈德斯坦(David Goldstein)检测了中国大陆和台湾携带M122的人群中的一个被命名为M119的微卫星的多样性,发现其出现频率非常高。但是,在马来西亚半岛和印度尼西亚地区,这个M119的频率却开始显著降低了。这个形态精确地表明一万年前的人口扩张浪潮的起源在中国,这与水稻农业的考古学证据完全一致。
M119和M122都是M175的后裔,东南亚地区人群的Y染色体上同时携带M119与M122的人口超过50%。对比之下,欧洲携带新石器时代移民的比例仅为20%。所以,东亚发生的移民不是浪潮,而是海啸。
人口大量增加更加适于发展农业,农业提供的稳定食物来源又导致了人口继续增加,所以,人们曾经认为,农业的效应完全是正面的。但是,各种学科的研究证实,农业的成就可能被过度夸大了,历史真相是农业带来了很多负面效应。
第一,农业时代的人均资源,远远低于狩猎采集时代,农业使得人类大大降低了抗击自然风险的能力。因为人类无法控制气候,而气候始终完全控制着人类。现在出土的证据表明,农业发展反而使得气候对人类的控制力更加强大。仙女木时期的农业人群可能经历了一段极其艰难的生活,但是继续狩猎采集的人群却没有受到影响。
仙女木时期(Dryas periods)系指冰河期结束时的一次反复,长达1千多年(1 300 ± 70年),气温下降8-20℃。1.28万——1.15万年前称为新仙女木时期(Younger Dryas),在此之前约1 000年还发生过一次旧仙女木时期(Older Dryas),全球气温突然下降,时间长达300年左右。这次气温骤降,给新石器时代从事农业的的人类带来很多困难和问题。
第二,农业人口的聚集生活带来的副产品是疾病的大量出现。
很多疾病必须聚集一定的人群数量才可能发生和传播,并且限定在某一区域内传播。例如,天花和伤寒必须在数万人的范围内才能传播,并且限定在这个范围内。人们过去认为,狩猎采集的人群比较易于受伤,所以寿命比较短,但是出土的骨骼证据表明,当时农业群体的寿命反而比狩猎采集的群体更短,原因很可能就是疾病的流行。
农业首先驯化了植物,随后驯化了动物,大量动物聚集生活也会产生疾病,再传染给人类。例如,人类的麻疹与牛瘟病毒就密切相关。威廉姆·麦克尼尔(William McNeill)认为,《圣经》中描述的多种瘟疫大流行正是农业传播到欧亚大陆时期,人类感染各种传染病的忠实记载。
第三,农业时代导致社会的分化和阶层的出现。
一般来说,狩猎采集群体中人与人是平等的,没有社会分工,现在桑人(San)部落和澳大利亚土著依然如此。农耕时代,初期阶段(田园牧歌时期)的大型战争比较少,人口增长很快,财富积累和社会分工出现,最后不可避免地出现社会分化,形成阶层,随后出现权力争夺和帝国雏形,引发前所未见的巨大规模的战争和破坏,导致人口多次大量死亡。人口减少反过来又造成大量土地荒芜和疾病流行……这种恶性循环,在各地的农业社会多次发生。
既然农业具备这么多负面效应,为什么人类还是喜欢农业?
事实并非如此。在世界各个角落都有排斥采用农耕生活方式的群体继续存在,他们的环境至今保护较好。但是世界大部分地区还是采用了农耕生活方式,并且不再回头。因为谁都不愿意天天拿着武器,出去寻找下一顿晚餐。
现在的研究已经证实,导致人类死亡的主要疾病的原因,分为三个阶段。
一、在狩猎采集时代,主要的死因是外伤,尤其是狩猎活动中的伤亡。
二、在农业时代,人类的主要致命疾病是传染病,尤其是源自动物的疾病,如天花、伤寒、霍乱、麻疹、肺结核、流感、黑死病等都与动物有关。在抗生素和疫苗发明以后,大部分传染病得到控制或消灭。
三、在现代,非传染慢性病是人类的主要疾病,例如高血脂、高血压、糖尿病、心脏病、脑血管疾病等。这些疾病都要终生服药,且都与基因有关,已经与传统的传染病的概念完全不同。癌症的起因也是基因突变的累积结果。
但是,某些传染病现在仍然存在,例如疟疾。
1 86 1年,法国探险家亨利·穆奥(H enri Mouhot,1826-1861)因为疟疾死在老挝的丛林里。亨利·穆奥在泰国、柬埔寨和老挝探险3年并发现了吴哥窟。虽然当地的居民知道吴哥窟,但是,是亨利·穆奥死后才发表的著作《暹罗、柬埔寨和老挝游记》(Travelsin Siam, Cambodia and Laos)使西方读者第一次知道了这座古城。
吴哥窟建筑群是东南亚当时最强大的高棉帝国的统治者在9-15世纪期间建造的,该建筑占地1 000平方千米。这是产业革命之前世界最大的城市,其巅峰时代,容纳的人口超过75万,但是在15世纪却被放弃了,除了吴哥窟寺庙(Angkor Wat)外,其他寺庙和建筑都被丛林掩埋。
这座城市为什么会被放弃?
假设很多,后来比较被公认的一个原因是生态压力。支持75万人口必需的资源之一是水源,当时只能来自周围的河流。14-17世纪,北半球气候变化,导致东南亚季风改变,由于缺少降雨和水源枯竭,人类被迫放弃水稻种植。这是吴哥城的设计者始料不及的天灾。
另外一个原因是疟疾。疟疾(malaria)是意大利语,意思是“坏空气”。疟原虫在污染环境里才会大量繁殖,通过蚊子叮咬传染人类。意大利人起的这个名字是正确的,疟原虫有两个宿主:人类和疟蚊。传播疟原虫的疟蚊(Anopheles,又称按蚊)有40多种,分布在世界各地,大部分疟蚊是热带和亚热带品种,大多在非洲、东南亚和拉丁美洲。疟原虫可以在疟蚊体内生活,也可以在人类的红细胞里生活。无处不在的疟蚊传播了疟疾,很多史前历史学家猜测,疟蚊曾经在人类进化史上给人类带来了毁灭性的灾难。
1992年,法国流行病学家雅克·维德拉格(Jacques Verdrager)发表文章认为,放弃水稻种植后,生态环境恶化,疟疾流行带来死亡,导致更多人放弃水稻种植,疟疾更加流行,死亡更多……仅仅几代之后,吴哥城就成为一座空城。疟疾在这座巨大的古城沦为废墟的过程中,扮演了关键角色。
疟疾的故事并未到此结束。
在1992年,DNA技术尚未成熟。随着技术的成熟,DNA密码揭示了疟原虫的秘密。
美国国家健康研究院(National Institutes of Health)的迪尔德丽·乔伊(Deirdre Joy)和她的同事发现,疟原虫在5万年前开始多样化,这个时间恰好是人类走出非洲的时期,暗示人类带着疟原虫前往世界各地。乔伊还发现了其他证据,一万年前,疟原虫开始大规模的多样化,这个时间正是新石器革命的农业起源的时间。
另一项DNA研究佐证了上述发现。宾夕法尼亚大学的遗传学家莎拉·提什科夫(Sarah Tishkoff)和她的同事仔细分析人类基因组上围绕G6PD基因的遗传变异,他们发现,一万年前,G6PD曾经受到强大的自然选择压力。
G6PD是细胞里的一种酶,可以把葡萄糖转化成一种亚细胞能量包(subcellular energy packet),这种亚细胞能量包名为NADPH,是人类细胞能量活力的来源。我们吃下的谷物——碳水化合物又称多糖类,被转化为单糖(葡萄糖)后,最终变成我们细胞里的三种能量:NADPH、NADH和ATP。所以G6PD极其重要。
我们体内的G6PD的精细调制的进化历史,已经长达几亿年,属于最重要的酶之一。在人类的身体里,如果G6PD缺乏或功能减弱,就会出现类似蚕豆中毒的症状:贫血、黄疸、肾脏失调……在红血球里,G6PD非常活跃,疟原虫也是在红血球里非常活跃,不知道什么时候,疟原虫发生了基因组变异,它们可以通过“虹吸原理”吸收G6PD的能量,从而成为红血球里的寄生微生物,直接破坏红血球的新陈代谢,最后迫使红血球“自杀”,同时也杀死了自己。
为了查清疟原虫的基因什么时候发生了这种变异,莎拉·提什科夫和她的同事对G6PD基因的变异进行了计算分析,发现变异出现了两次:
第一次,3 840-11 760年前,主要发生在非洲。
第二次,1 600-6 640年前,主要发生在地中海。
G6PD基因的这两次变异,都发生在农业出现之后。这是一个令人震惊的发现,结合疟原虫的基因变异,证明在过去的一万年里,疟疾已经成为人类的主要噩梦之一。
疟疾原本是狩猎采集时代的一种古老疾病,后果并不严重。在定居和农业开始后,疟疾的威胁日益增大。中东地区水利灌溉为疟蚊繁殖创造了机会,吴哥窟周围的稻田水塘湿地也是疟蚊繁殖的好地方。
农业,迫使人类基因组变化了,也迫使致病微生物的基因组变化了。
农业,引发了基因与基因的战争。现代的飞机,为这些涉及基因层次的疾病传播提供了远远超过远古步行的人类之间的传播速度——疟蚊带着疟原虫,可以乘坐飞机很快到达其他大洲。这种坐飞机传播的涉及基因层次的疾病还有克雅二氏病(Creutzfeldt-Jakob)和艾滋病(HIV)等。
农业,就像一把双刃的镰刀。
不能与上帝开玩笑
历史上,只有一个国家做过制造人类的实验,苏联。
1924年,伊万诺夫(Ilya Ivanovich Ivanov,1870- 1932)向苏联政府写信,希望开展人与灵长目动物的杂交实验。列宁的秘书戈布诺夫(Nikolai Gorbunov)对此很感兴趣,批准了这个实验。
1927年,伊万诺夫教授来到几内亚,给两个雌性黑猩猩做了人工授精。两个雌性黑猩猩都没有怀孕。没有人知道精子来自什么人,但是知道陪同教授的是他的儿子。伊万诺夫又来到刚果,声称要在医院里实验,但是没有说什么实验。当地的总督坚持要在室外观看实验,伊万诺夫拒不同意。实验在两个非洲国家都失败了。
斯大林的故乡,格鲁吉亚的苏呼米(Sukhumi)有一个灵长目实验站表示欢迎教授。伊万诺夫教授从非洲带回几只黑猩猩,包括一只雄性黑猩猩,来到苏联领袖的故乡准备继续实验。他改变了办法——用雄性黑猩猩和苏联女性杂交,可能成本低一些,而给雌性黑猩猩授精的难度比较大。
1928年,伊万诺夫开始寻找志愿者。他需要5个苏联妇女为科学献身,但最终只来了一个志愿者。苏联档案里的这个女人的代号是G,不知道G的年龄多大,是哪里人,长得漂亮还是丑陋。苏联档案里只有G写的一封信:“亲爱的教授:我的私生活会被毁灭,我将看不到继续存在的意义——但是想到能为科学服务,我还是鼓起勇气给你写信。我请求你,不要拒绝我。”
教授没有拒绝她。
伊万诺夫和G做了一系列准备,要在苏呼米为G授精。不幸的是,由于缺乏食物和牛奶,这只唯一的雄性黑猩猩死去了。
1930年,伊万诺夫教授被捕。
1932年,伊万诺夫教授在集中营里饿死了。
1938年,列宁的秘书戈布诺夫也被处决。
但是,正常的“制造人类”的实验并非不存在。各种原因的不孕症是困扰人类多年的难题,剑桥大学的罗伯特·爱德华兹(Sir Robert Edwards,1925-)从1960年代开始研究人工授精的可能性。
1968年,世界第一个体外试管受精成功。
1978年,世界第一个试管婴儿路易斯·布朗(Louise Brown)诞生,她母亲患有输卵管阻塞,9年无法成功怀孕。这一技术现称IVF,即体外受精。
2010年,爱德华兹获得诺贝尔奖,这一年世界体外受精婴儿突破400万人。需要说明的是,爱德华兹的技术并不涉及基因。
另一个英国医生走得更远,他试图“设计优秀的人”,他要挑战基因。
2002年,世界第一个“设计婴儿”(designer baby)在英国诞生。这个婴儿查理(Charlie)成为媒体头条新闻。起初查理似乎很健康,3个月后出现明显不正常,被确诊为DBA贫血症(Diamond-Blackfan anemia,没有正式中文译名),这是一种在整个英国也找不出几个的罕见疾病:红血细胞先天不足,无法携带足够的氧。各种减轻症状的治疗方案都失败了,查理可能死亡,除非找到一个合适的骨髓捐赠者替换查理的造血骨髓。但是,寻找合适骨髓捐赠者的种种努力也失败了。
如果查理有兄弟姐妹,可能骨髓适合,但是查理是唯一的孩子。于是,唯一的希望是查理的母亲再生一个孩子,给查理捐献骨髓。这个方案也有风险,因为查理父母的第二个孩子也非常可能患DBA贫血症,所以查理的母亲必须找其他男人授精生孩子,这已经不是医学范畴而是伦理道德范畴的问题了。
查理的医生穆罕默德·塔拉尼西(Mohamed Taranissi)曾经是一个非常成功的医生,他原本希望“设计婴儿”成功并推广这种技术。他现在进退两难,饱受媒体和舆论的猛烈攻击。
英国的法律不允许这种授精,美国的法律允许。于是,塔拉尼西飞到美国芝加哥寻求帮助,13个人愿意捐精。塔拉尼西自己掏腰包承担全部费用,请查理的父母也飞到芝加哥,在13个人中选择合适对象。最后选择了两个人的精子,一次性全部植入查理母亲的子宫。几个星期后,超声波检测证实查理的母亲怀孕了,只怀了一个。
18个星期之后,抽羊水检查确认怀孕婴儿的骨髓适合移植给查理,所有人都松了一口气。但是10个月后,发现婴儿是臀位分娩,于是查理的母亲不得不施行剖腹产,生下第二个儿子杰米(Jamie)。这位30岁的英国妇女始终非常合作,她看着5岁儿子查理的病况,“心都要碎了”。
常年服药的查理,又开始了化疗。他的免疫系统被摧毁,体重减轻,头发几乎掉光。随后查理接受了骨髓移植,并大量服用防止排异反应的药物。6个月后查理停止了服药,多次检查后证实,查理的红血细胞正常了。
10年后,查理的父亲说了一句话——“我们不能和上帝开玩笑”。
语言造就了人类
1996年,伦敦儿童健康研究所(Institute for Child Health)的一群医生迎来了牛津大学遗传学教授安东尼·摩纳哥(Anthony Monaco,1959-),他们要会诊一种奇怪的疾病。医生们发现,一个源自巴基斯坦家族的几家亲戚,三代人都罹患了语言障碍的遗传病。为了保护隐私,这些家族被称为KE家族。他们无法控制面孔下半部的肌肉运动,因而丧失了语言表达能力,他们也无法理解别人的语言。
安东尼·摩纳哥教授进行了基因组扫描(genome scan),分析对比几百处可能发生变异的位置,包括KE家族中患病和没有患病的成员。经过一年多的努力,发现问题可能出在7号染色体,但是却无法确认是哪一个基因造成的。
幸运的是,后来牛津大学又遇到了另外一个独立的患者,代号CS,这是一个与KE家族无关的男孩,也罹患了类似的语言障碍。摩纳哥和他的同事们再次进行了基因组分析,发现这个男孩出现了染色体的重新排列,染色体的某一部分分裂开了,分叉伸进另一个染色体里,这种现象被称为基因置换(translocation)。在这个分叉点上,有一个基因的功能因此紊乱了。这个男孩CS的这个突变,在KE家族的染色体上也找到了。
这是人类第一次发现影响语言的单一基因,这个基因被命名为FOXP2。
2001年,《自然》(Nature)刊登出这个发现后,引起巨大轰动,有些媒体报道为“发现语言基因”。
FOXP2负责制造带箭头分叉的P2蛋白,这类蛋白传递很多DNA的打开与关闭信号,所以又被称为“基因组的分子公共汽车”。这些DNA涉及体内的物理部分和精神部分,所以控制了语言和语法等,在此之前,人们曾经以为是几百个基因在操控语言。
FOXP2的基因调控中枢角色,在黑猩猩和老鼠身上也得到了发现和证实。老鼠的这个基因发育不全,类似婴儿。所以FOXP2的进化应该超过7 000万年。人们立刻联想到,FOXP2在人类的语言进化中必然扮演了重要角色,南猿、能人、直立人和尼安德特人,可能都有发音沟通的能力。
在以色列的科巴拉洞穴(Kebara Cave)出土了几乎完整的6万年前的尼安德特人的骨骼,包括完好无损的舌骨(hyoid)。舌骨是娇嫩精巧的骨头,可以帮助人类说话。尼安德特人也有舌骨,表示它们也可能有说话能力。
2007年,斯万特·帕博(Svante Paabo,1955-)的团队发表了他们极其惊人的研究成果:尼安德特人的FOXP2基因,与人类没有什么差异,也就是说,尼安德特人与人类一样具备语言能力。(斯万特·帕博的团队从克罗地亚北部的文迪迦洞穴(Vindija Cave)获得了尼安德特人的FOXP2基因。这个洞穴的堆积层的年代为2.5万——4.5万年,最下面是尼安德特人的遗骸,上面是现代人的遗骸)
帕博的团队研究了5年,他们发现人类和黑猩猩的FOXP2基因的蛋白上都有715个氨基酸,其中只有2个氨基酸的序列不同,这一差别可能出现在500万年前。但是,尼安德特人的FOXP2基因,与以前的猜测完全不同,确实具备说话的能力。但具备说话能力,并不意味着拥有语言。很多鸟类、动物和鲸鱼都有发音沟通的能力,甚至青蛙和昆虫也可以做到通过声音沟通。科学研究证实,黑猩猩即使经过训练,最多只能表达1-2个单词。
这个“小小”差异,意味着什么呢?举一个简单的例子。“妈妈从冰箱里取出苹果”这个句子包含全部信息,既有地点,也有先后次序,还有相应动作。对比之下,妈妈冰箱,冰箱苹果,妈妈苹果,妈妈取……表达1-2个单词没有任何意义。
语言,正是“亚当”“夏娃”的苹果,使得人类区别于其他生物。
语言的出现,使得人类成为人类。
不论什么原因,6万年前走出非洲的人类,在世界所有角落都留下了抽象思维和想象力的石刻证明,标志着艺术从开始就伴随着人类生活。创新是人类的能力。创新是一个复杂的过程,创新的核心是思考和实施解决某一问题的抽象思维,创新的第一步是想象力。艺术是想象力和抽象思维的产物——只有语言才能交流这些看不见的想象力和抽象思维。
语言、艺术创造了人类。人类创造了人类自己。
设想一下,每天晚上,在远古的篝火边,大家围着火堆讨论当天的故事和先祖的传说,雕刻着艺术作品,如何捕捞三文鱼、猎杀猛犸象,如何攻击狮子?如何在洞穴绘制壁画,如何在岩石上雕刻浮雕?他们也会议论其他部落的新闻……
这不是凭空的想象,这是仍然保留在现代的世界各地的游牧民族的日常生活……
信仰、图腾、宗教的诞生,全部因为语言。
他们是米开朗基罗、莎士比亚和爱因斯坦的祖先,他们在亚欧大陆的无边无际的广袤的干草原上自由驰骋,没有任何约束和限制。
现代智人的考古记录,事实上正是一部人类创新的历史记录——他们迅速适应了各地的气候和环境,开始尝试驯化小麦、水稻、玉米,虽然这些创新遭遇过多次失败和失误。狩猎采集时代的语言多样性、文化适应性和无拘无束的创新性,曾经是所有多元文化的源头。在美洲的猛犸象的尸体里,曾发现十几个美洲土著的石器枪头。在中东和希腊地区发现很多人类主动攻击狮子的浮雕……我们的先祖,曾经几乎无所不为,为所欲为。
进入农业社会之后,人口数量和密度不断增大,出现了太多有形的和无形的“边境线”,既约束了自由也约束了思想。大大小小的王国和帝国、印度的种姓划分、天主教的等级森严、孔儒的停滞僵化、欧洲的封建体系等,都曾经使社会长期停滞不前,不再出现创新。
正在消失的语言与文化
封闭的苏联时代隔绝了遗传学,苏联时期的哈萨克斯坦、乌兹别克斯坦等地区成为基因形态的“黑盒子”。苏联解体后,美国遗传学家们赶紧前往“黑匣子”地区给少数民族采样,其中一个少数民族是亚诺比人(Yaghnobi people)。亚诺比人说亚诺比语(Yagnobi),他们的历史可以直接上溯到丝绸之路。
1 500年前的亚洲中部地区,从波斯到中国的丝绸之路上的所有贸易中心,曾经都说粟特语(Sogdian)。公元6-8世纪,穆斯林逐步征服这一带之后,粟特语类开始衰落。20世纪,粟特语系的大部分语言全部消亡,仅仅留下亚诺比这一种语言。
现在的亚诺比人居住在塔吉克斯坦北部遥远的扎拉夫尚河谷(Zarafshan Valley)的几个村庄里,他们是继续使用这种1 500年前的古老语言的仅剩的小小群体。美国遗传学家计划拜访他们,说服他们合作提供DNA的血样,追踪历史的遗迹。
走过几个平行的河谷,又在崎岖坎坷的道路上跋涉了几个小时,美国人才发现一个村庄。可是那里的老人却说他们这一趟白跑了。老人解释说,亚诺比人世代居住在这里,可能丝绸之路时代就住在这里。1960年代开始,因为旱灾严重,苏联政府把他们迁移到低地。1980年代,这一带发生地震,幸存的亚诺比人都搬到首都杜尚别(Dushanbe)居住。在这片故土上已经找不到亚诺比人了,他们都在首都当清洁工或卡车司机。不过老人也给了他们一个希望,距离这里几天路程的山区里,可能还有一个亚诺比人的村庄……
科学家们又进行了几天的艰难搜索,终于碰到一个偏远的亚诺比人的村庄,人们非常乐意帮助这些遗传学家,带领他们在首都杜尚别一带寻找同乡。科学家采集到了出乎预料的大量血样,丝绸之路上一个与世隔绝的文化残迹出现了。
那位塔吉克斯坦的乡下老人讲述的故事,每天都在世界上发生。亚诺比人的故事,世界处处都在上演。一座又一座冒出来的现代化城市,吞没了一批又一批村庄,原始居民和语言都湮灭了。有的社区对多样性非常宽容,有的社区认为多样性是统一的障碍。新生的政权总是努力推进语言的统一性。
让我们看一看19世纪的欧洲的语言的真相。
如果访问今天的法国,游客们都会对法国人热爱法语印象深刻。国家语言的官方捍卫者法兰西学术院(Academie francaise),像猎鹰一样紧紧监视着说的法语、写的法语,以免受到国外的“不良影响”。
其实,仅仅150年前,大约6代人之前,法国领土上说“法语”的人口还不足一半,大部分“法国人”说自己当地的方言,甚至自己的语言。
同一时代的意大利,说意大利语的人口不到10%。奥地利首相梅特涅(Clemens von Metternich,1773-1859)说过:“意大利不是一个国家,只是一种‘地理表述’。”如果不算语言,梅特涅显然说的不错。
19世纪的欧洲,出现了各种新思想和运动,浪漫主义、现实主义、工业化、殖民扩张都对“现代”的世界观发展产生了巨大影响。其中最重要的思想之一是民族主义,不仅造就了现代欧洲的政治版图,并且流传到世界各地。
19世纪之前,欧洲分裂为各自独立的封地采邑——王国的领地和公爵的领地。人们的生活更有“区域性”,仅仅与周围发生的事件相关,欧洲历史上的配偶们的出生地之间的距离没有多少千米。这种特点,造成了语言的延续性。
现代法语是法兰西学术院捍卫的官方语言,但是在18世纪之前,法国存在的语言五花八门,分别可以上溯到各个省的地方语言,包括巴斯克语(Basque)、布列塔尼语(Breton)、奥克西坦语(Occitan)、科西嘉语(Corsican)、阿尔萨斯语(Alsatian)等完全不同的语种。民族主义流行欧洲之后,各个政府为了寻求文化的统一性而偏好某种一语言,不喜欢另外一种语言。英国政府喜欢英语,于是支持英语,贬损凯尔特语族。19世纪,统治者开始以民族语言进行强制性的学校教育和军队服务,仅仅几代人就完成了一种语言的传播。
德语的诞生,更是出于精心的设计。几乎全世界的孩子都知道《格林兄弟童话》,但是却不知道格林兄弟是为了通过童话传播“正规的德语”。雅各布·格林(Jacob Grimm,1785-1863)是语言学家,他制定德语的发音规则,维护和铸造德语的民族同一性,他是“德国民族”的精明建筑师之一。
现在,15种最大的语言的使用人数占世界人口的一半,前100种语言的人数占全球人口的90%以上。但是在公元1500年,估计存在1.5万种以上的语言。形成一种语言需要500-1 000年的时间,每一种语言承载着一种文化。现在,全世界仅剩约6 000种语言,平均两星期消失一种语言,21世纪末将再消失一半。
英伦群岛的原始土著的语言之一是凯尔特语族的马恩岛语(Celtic Manx language),1874年有1.2万人说马恩岛语,20世纪之交只剩4 000人,1974年最后一个说马恩岛语的人死去之后,马恩岛语成为活化石,现在仅仅剩下几百个爱好者了。
罗曼语(Romance languages)源自拉丁语,随着罗马帝国的兴盛传遍欧洲,今天的法语、西班牙语、意大利语、罗马尼亚语、加泰罗尼亚语(Catalan,西班牙官方语言之一)、罗曼什语(Romansch,瑞士官方语言之一)的先祖之一都是罗曼语。
语言记录了历史,一种语言的消失就是一种文化传承的断绝。
基因的分析研究,有时必需语言和文化的帮助。原因有二:第一点,我们的先祖生活在五万年前,仅仅经历了大约2 000代,突变的产生并不频繁。我们只要做不多的采样和大量的分析计算,就能找到某一个点位的大部分多样性,尤其是Y染色体多态性比较丰富的部位。由于人类的“种族”太年轻了,物理化石差异不大,大部分非洲、亚洲和欧洲的化石都很相似,无法分析推测出他们的肤色、发型和其他外貌特征。遗骨的证据证明,我们所谓的种族概念是一种非常晚近的现象,直到最后一次冰河期结束之后,现代人类才开始“发散”出现在看到的多样形态。
第二点,人类是机动的,各个群体在历史上是一边迁移一边混血,所以相互之间的遗传变异并不明显。即使我们走出非洲之后,出现了各种遗传基因标记,也被广泛散布。语言消失的动态加速,表明人类的混血也在加速。很多语言正在消亡,表明原先互相隔绝分别生存的群体,正在融合为更大的群体。
那么,是否存在统计这些现象的具体数据?
美国的人口统计就是一个典型的例子。2000年统计的美国的人口是2.814亿人,比1990年增加了13%,并且显示出更多的民族变化版图——美国的人口统计的“种族类型”从5类增加为63类。申报是“白人与少数民族混血”的后裔的人数,增加到680万人。真实的混血的类型、混血人口的比例,估计比政府统计数据更多更大。有些人已经不知道应该归属为哪一个类型。比如,人人都知道的高尔夫球手“老虎”伍兹(Tiger Woods),但是他自己不知道自己属于哪个“种族”,他的祖先包括非洲人、美洲人、欧洲人、东南亚人……美国越来越成为“种族”的熔炉。这种融合可能是一件好事,混血的下一代比他们的前一代或许拥有更多的优点。
产业革命造成的机动性,导致人类历史的第三次大迁移,也带给我们一幅全新的遗传画面。五彩缤纷的多样性,可以把五万年前开始分离的人类识别出来,并再次分门别类,现在发生的全球性种族大融合,则是人类历史上过去从未发生的新现象。
现代化的浪潮使城市正在吞没乡村,孤单生活的群体正融入难以计数的人海。虽然基因标记无法消失,但是同时也融入了无边无际的人海……于是,他们携带的基因故事同时化为更加难解的历史谜团。
我们必须知道我们的过去,才能预知我们的未来。
我们终于具备了这种能力,因此我们也被赋予了一种责任——这是人类的责任,这是我们从非洲出发殖民全球必须承担的责任。
第七章 农业文化的反思
大约20万年前,现代人从非洲开始。
大约6万年前,现代人离开非洲,走向世界。
今天,我们生活在一个高度全球化的世界,一个人与其他人之间发生联系的方式超过一个世纪之前的想象。非洲人、欧洲人、亚洲人、美洲人……正在再次融为全球性的一个混合群体,这种混血现象和融合程度是人类历史上的第一次。在DNA的层次上,我们都是一样的,我们都是日益扩大的人类大家族的一部分。这个世界将因为技术进步日益加速的节奏,在未来几代人的时间里发生超乎预期的改变。
我们现在日常生活中的一切,大部分是最近几十年才进入每一个家庭的。
E-mail、Google、手机、电脑、混合动力汽车……以互联网为核心的全球性社会网络已经成为人们须臾不可分离的生活组成部分,离开这一切已经不能想象。但是,现代生活方式也成为人类的巨大包袱。越来越多的非传染性慢性病,如高血压、心脏病、糖尿病、高血脂、肥胖症、癌症……正在不断增多;各种精神疾病、心理失调、沮丧、焦躁、忧虑……也在不断增长。
的确,我们是自己命运的主宰,但是,究竟是我们创造了农业文化,还是这种文化驱动着我们?
一万年前的农业文明,彻底颠覆了我们的田园牧歌。越来越多的证据使人们逐渐发现:正是农业文化的出现,开始损害人类的健康。
考古证据证明,转向农业生活的动机决定了今天的复杂世界的出现,从工业革命到互联网到生物时代,人类稀里糊涂冲进了21世纪。但是,世界各地的人们仍然在没有止境地开发土地,生产更多食物的强烈欲望冲击着大自然设定的底线。
我们这个物种具备制造各种恶果的能力,我们已经并在继续导致更多其他物种的灭绝。
遗传学和DNA迫使我们重新认识,过度贪婪的农业文化带来过度需求导致的一系列后果。
巨大的进化压力
20世纪,人类发现基因是遗传变异的来源,开始研究基因。
1987年,25年前,在人类的技术可以观察DNA之后,全世界的注意力都转向了DNA。人们发现,在几十亿年的进化中,各种生命并非“互相战斗”而是“互相合作”地占据了地球,共同演化出了一个绚丽多彩的大千世界。在50多年的DNA研究过程中诞生了几十位诺贝尔奖得主,达尔文似乎被淡忘了,“适者生存”的自然选择似乎被“中性理论”取代了。
2007年,《PLoS生物》(PLoS Biology)发布了芝加哥大学乔纳森·普里查德(Jonathan Pritchard)研究小组的一项研究结果:达尔文的“自然选择,适者生存”理论不仅是正确的,并且在最近一万年里对人类的基因组产生了重大影响。也就是说,人类的很大一部分“进化”,是在农业出现后发生的。
那么,人类在这一万年里,为什么承担了如此巨大的进化压力?更直白地说,为什么我们取得如此巨大的物质进步的同时,每一个人都感到压力巨大?甚至觉得物质满足并未带来应有的幸福感觉,并且不得不随着继续的快速发展而“进化”自己身体的各个部分?
这是最新的DNA研究向全世界提出的一个问题。
芝加哥大学的这项研究的基础数据,来自2005年启动的“国际人类基因组单体型图计划”(International HapMap Project)。与群体遗传学的单倍群不同,单体型(Haplotype,希腊语原意单一,单体)研究每一个个体的微小的遗传变异——单核苷酸的多态性(Single-nucleotide polymorphism)。这篇论文,甚至描述了我们的眼睛、耳朵的精致进化演变的原因和过程。
最近一万年里,人类的环境和演变,确实过于巨大了。
普里查德用珠子项链为例解释自己的发现。
每一个人的单体型,就像这个人自己佩戴的珠子项链。一个长达2米的基因组被切分成23对46个染色体小股,染色体DNA序列就像一串串珠子将它们串联起来。在结合和重组时,每一代人的父母的珠子都被拆开打乱,按照另外一种顺序重新串起——这些珠子形成一个新的项链。下一代携带的新项链,与父母的项链都不一样,他们的再下一代的项链,又是另外一种新的顺序的珠子项链。经过若干代之后的项链,已经面目全非——父母原来的项链已经湮灭了,消失了。于是,形成了单体型的多样性。
在基因序列里,只有线粒体DNA和Y染色体DNA不参与重组,所以根据它们可以统计分析计算出人类先祖的踪迹,找到人类的旅程。
图片出自韦尔斯《潘多拉的种子》
现在,其他的22对染色体的单体型的更加复杂的多样性,也得出了同样的分析结果。
普里查德领导的小组开发出一套新方法检测基因组中的单体型,他们的样本来自欧美、非洲、亚洲等地。每一个人的项链都不一样,所以每一个人都不一样。这些项链就像某种“遗传时髦”,根据气候和环境而发生形态的变异。这是大自然的力量推动的,具体的原因还不清楚。
普里查德应用的方法属于集成单体型积分(integrated haplotype score, IHS),很多微小的变异,积累到一定程度就会成为变异,甚至非常显著的变异。过去人类发生过多次显著的变化,例如拇指与四指分开、肤色改变、大脑神奇发育……这些当年的“时髦流行”成为人类与其他物种的区别。
在每一个人和其他人之间,也存在差异,因为单体型的形态各不相同。在23对染色体上,普里查德领导的小组发现了几百个基因区域,分布相当稀疏,受到某种自然选择的强大压力,甚至影响到人类不同群体的眼睛和耳朵的精细区别。
最令人震惊的是,这些变异就发生在最近一万年里。
人们过去认为,这种变异必须花费很长时间,必须经过很多世代。这个小组发现,这些变异竟然如此年轻。人类的单体型的形态,发生如此显著的变化——高度适应各种环境的生物组织变异,仅仅从大约一万年前开始,仅仅经历了大约350代人。这是一种什么样的巨大选择压力,导致基因组出现如此显著变化?
普里查德领导的小组,最初也不敢相信这一事实,但是经过多种验证核实证明,这一切都是真实的。
这意味着什么呢? 根据已知的考古地点以及基因组的线索,估计在20万——8万年前,我们这个物种的数量不多并且相对稳定,因为化石数量很少。即使12万年前出现在南非和中东,人数也没有显著增多,中东只是非洲的地理延伸,气候、植被和动物也类似,这些小小的群体只是四处游荡,没有冒险离开非洲的故乡。
从曲线上看,10万年前,人数只有几万人。在8万——5万年前的冰河期时代,不知道什么原因,人类化石证据出土很少,包括非洲也是如此。
我们人类可能因为某些灾难性的挑战而撤退了,人口数量急骤下降。根据遗传科学的对比推算,当时人类比大猩猩的数量少,7万年前只有2 000多人,濒临灭绝的边缘。6万年前曲线开始上升,人数增长并开始走出非洲。此后经过4.5万年,人类散布到世界所有大洲(除了南极洲),从几千人变成几百万人的狩猎采集者。
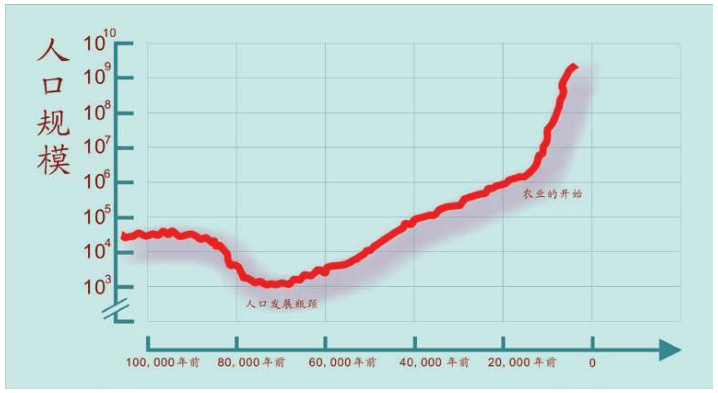
真实比例的人口爆炸示意图
大约一万年前,出现一场重大革命。人类定居下来,做出一个改变与大自然关系的重大决定,这就是发展农业。曲线上的人口开始快速增长,由几百万人变成70亿人,这场人口大爆炸的结果是人类统治了世界。
狩猎采集者依赖食物资源,农业人创造食物资源。这个动机改变了历史。控制了食物的来源,就拥有了在特定地点选择养活多少人的能力。
普里查德的基因研究结果显示:最显著的功能形态变化是皮肤的黑色素的变化,五种不同的基因涉及欧洲人的黑色素的选择,其中作用最强的基因之一是乳糖分解酵素(Lactase)。乳糖分解酵素帮助人类代谢乳糖,否则就会出现乳糖不耐受症。这个基因在人类婴幼儿时代启动,帮助消化母乳,此后大部分群体的这个基因关闭,成人不宜消化乳糖。
1万——0.8万年前,中东人驯化了山羊和奶牛,增加了营养来源。这些中东人把山羊和奶牛带到欧洲,长期饮用乳品造成一个基因突变,现在90%以上的欧洲人把乳品作为日常饮用品,但是亚洲和非洲的群体中的成人,继续呈现对乳糖的不耐受症。
这个例子诠释了遗传中自然选择的作用:欧洲成人也分泌乳糖分解酵素。
普里查德的技术特点是“不做任何假设”地分析,仅仅从海量数据中查找是否存在自然选择的痕迹。这是人类基因组工程的重大成果。过去,我们必须艰难地采集和烦琐地分析少量的基因序列数据,以证实某一种假设。现在,情况完全颠倒过来了,基因序列数据成为洪水,我们必须对无数分布形态的统计学分析结果作出合理的解释。
普里查德领导的小组还发现,人类的一个负责酒精脱氢酶(alcohol dehydrogenase)的基因,也受到自然选择的压力,这个基因产生的酒精脱氢酶可以分解进入人体的白酒和啤酒。有趣的是,他们还发现1号染色体上的一个基因,也受到自然选择的压力,导致肝脏组织发生变异,可以分解外来的化合物,例如进入身体内的各种药物。此外,随着各种新的食物进入我们的身体,我们也需要肝脏的这些“清洁”基因的功能,将这些新的食物中性化。
普里查德的最后一个发现是很多基因出现重叠(overlap),证明自然选择和基因的冲突引发了复杂的综合性疾病,如糖尿病、高血压等。例如,高血压只是一种症状,并非一种疾病。导致高血压的原因很多,这种“疾病”极其复杂,既涉及遗传,也与生活方式有关。其中一种高血压类型是“盐敏感”,这类高血压患者如果吃了一定数量的盐,就会强烈影响血压的升高。几乎一半高血压患者呈现出这种“盐敏感”,在1号染色体上的细胞色素P-450(cytochrome P-450)的一组基因CYP3A导致了这种“盐敏感”。基因的这种变异是保护性反应,阻止我们的生活方式发生剧烈改变——农业的高度发达使得我们可以成吨地吞进各种营养,但是,人体根本不需要存贮这么多营养。
普里查德的计算分析证实,从狩猎采集生活转向农业生活之后,我们的DNA受到了巨大的影响。达尔文发现的自然选择,不仅产生了很多正面变化,例如皮肤颜色变淡、可以消化牛奶等,也造成很多负面效应。农业发展促成了人口大爆炸,人口大爆炸使我们与过去决裂,但也留下了遗传学的不利后果。
人类的“进步神话”之一是:过去一万年,人类摆脱狩猎采集生活,进入物质极大丰富的时代。大部分人认为,我们的先祖此前的生存异常艰难,英国哲学家托马斯·霍布斯(Thomas Hobbes,1588-1679)形容他们的生活是“孤独的、贫穷的、危险的、野蛮的、短寿的”。所以,当农业和政府一起降临时,优越性似乎是明显的,人们的生活似乎可以无限制地改善下去,人口与食物的增长都是正面的。但是,考古证据发现的事实恰恰相反。
1984年,人类学家约翰·劳伦斯·安吉尔(John Lawrence Angel)分析了地中海东部地区的人类,在转换为农业生活方式之前和之后的各个时期的遗骸,尤其分析了牙齿(根据牙齿可以判断寿命),计算得出下表:
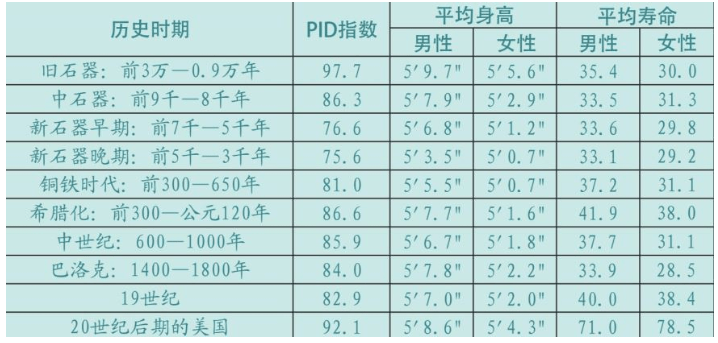
这些数据令人非常意外。旧石器时代人的寿命短,男性35.4岁、女性30.0岁的原因是婴幼儿死亡率较高;新石器时代开展农业后,人均寿命反而降低:男性33.6岁,女性29.8岁;中世纪身高降低,PID指数下降约22,此时的人类不是死于婴幼儿时期,而是死于疾病。
美国科学家也得出了类似的研究结果:农业生活方式,使得人类更不健康。
毫无疑问,如果不伴随着人口爆炸,农业肯定是拥有巨大效益的进步。我们曾经不断改良植物和动物的品种和产量——但是,我们是否更应该改良我们自己?如果我们再不控制土地资源的摄取和人口的增长,有可能像七万年前一样再次面对濒临灭绝的威胁。
农业新文化的成长
突尼斯城,曾经是迦太基帝国的首都。罗马帝国兴起于战胜迦太基帝国的三次长达120多年的战争。在突尼斯的克肯纳群岛(Kerkennah Islands),人们仍在沿用着几千年前的古老捕鱼方式:不是渔网,不是鱼钩,而是人造陷阱。
突尼斯人花费大约两个星期制作一个精巧的陷阱,放进地中海里,受骗的鱼类一旦进去就无法出来。这里的人们因此享受过几千年悠闲富足的海洋生活。
这种陷阱曾经非常有效,原来每天可以取出150磅活鱼,但是现在,每天进入陷阱的鱼类数量不到原来的十分之一。
地中海空了,人类的摇篮空了。
一万年前农业出现后,陆地的植物和动物食物来源,97%已被驯化了。
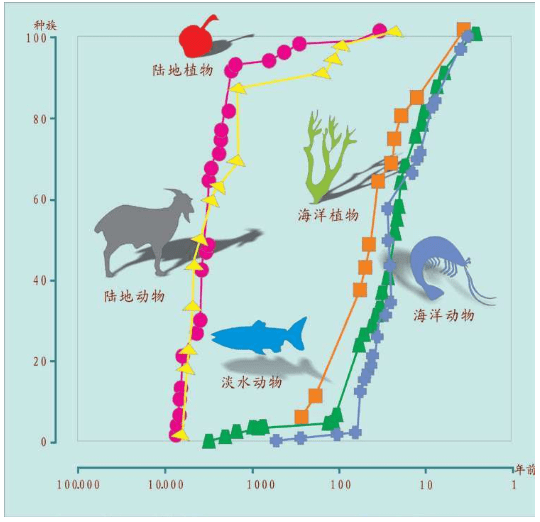
100年前,海上的植物和动物食物来源,在仅仅一个世纪内就被驯化了,而其中的四分之一是在上世纪最后10年这么短的时间内驯化的。
为了使得三文鱼的颜色在长期冷冻条件下依然显得栩栩如生,挪威的海上丰收公司(Marine Harvest)培育出可以长期保持鲜红颜色的新三文鱼品种。一万年前的农民,现在为海上驯化的新三文鱼品种申请了专利,进入了工业化生产。三文鱼曾经是捕捞最为困难的海上美味之一。这家挪威企业的驯化已经扩展到鳕鱼、比目鱼和金枪鱼,大量出口世界各国市场,中国市场上也充斥着这种“颜色鲜艳”的挪威三文鱼。
英国石油公司BP(British Petroleum)的业务遍布全球,从事海产养殖的子公司Nutreco在1994年与BP分离,现在员工近10万人,在30多个国家建立了100多个生产设施。他们在澳大利亚被称为“金枪鱼海上牛仔”。
中国驯化淡水鱼的历史超过2000年,曾经创造过科学循环利用废料的养鱼方式:宋朝驯化出青、草、鲢、鳙四大家鱼种类,并且将吐丝成绸的蚕的粪便作为鱼的饲料。20世纪后期开始,中国海洋水产驯化和人工养殖的范围已经遍布中国沿海地区。
欧洲天主教会曾经不允许吃肉,但是因为鱼类是“冷血”动物,教会认为不属于肉类,所以投资培育鱼类。欧洲首先驯化了海上的鳕鱼。这些技术现在传播到夏威夷和其他波利尼西亚群体——这些太平洋海岛上的人们也开始用鱼塘饲养鱼类,逐步取代了丰富的海产。
我们餐桌上的主要日常食物小麦、水稻、肉类、土豆……都是一万年前新石器时代后期的农民驯化陆地生物的成果。而海产驯化是由20世纪100年中海上的农民开始的,他们与先祖的区别在于拥有了高新科技和大规模投资。虽然世界的水产品中,现在只有大约四分之一是人工驯养的,但是人工驯养的比例正在不断提高。
捕鱼,是我们的先祖数百万年的狩猎采集时代遗留至今的唯一的生产方式。现在,过度捕捞的直接后果是世界海产品产量大幅度下降,1970-1980年是世界海产的巅峰,此后各种海产的收获全部出现崩溃。
1995年,鳕鱼的商业捕捞停止了。在英国的北海地区,鳕鱼被宣布为“商业性灭绝”了。继续徒劳地追寻残存的少量鳕鱼,已经无利可图。
2003年,全球29%的公海渔场“崩溃”,产量减少90%以上。
换句话说,世界的海洋和渔场衰败了。
戈登·柴尔德(Gordon Childe)把农业称为“新石器革命”,是人类这一物种的历史转折点。戈登·柴尔德认为,“人类自己创造了自己”,他勾画出冰河期结束时地中海沿岸气候变化和农业诞生的关系。他的证据和结论是基本正确的。拉尔夫·爱默生(Ralph Waldo Emerson,1803-1882)说:“第一个农民是第一个人,所有历史上的贵族都是拥有和使用土地的人。”
新石器革命带来很多负面效果。但是,人类一旦踏上这一步就不再回头,因为就如我们前文所说,谁也不愿意拿着武器,每天出去寻找下一顿晚餐。
具有讽刺意味的是,在贪婪的农业文化的驱动下,当海洋也开始衰败时,人们还在被反复警告:食用鱼类比较健康,鱼肉含有较少的饱和脂肪,尤其是鱼类含有omega3之类成分可以防治无数的“现代病”,从心脏病到老年痴呆症……所以,过去的30年里,欧洲和美国的猪肉和牛肉销量下降三分之一,水产品销量持续攀升。
于是,剩下的唯一出路就是人工饲养海产品。
我们先祖狩猎采集时代的唯一遗产——捕鱼可能很快将濒临灭绝,取而代之的是水产业:aquaculture,这个英文名词是“水上+文化”两个意思的合成。
六万年前人类走出非洲,随后很多大型动物灭绝了。捕杀大型动物才能收获更多食物。但是,中东的人类发现,定居种植草类和收集种子更可靠。
现在考古的方法之一是检测锶(strontium)的水平:遗骸的锶水平越高,人类食用植物的比例越大。中东的纳图夫人(Natufian)的锶水平曾显著增高,证明农业最先起源于中东。但是在1.28万——1.15万年前的新“仙女木时期”(Younger Dryas),全球的气温骤然下降8-20℃,时间持续1,300±70年,这段时期锶水平又显著降低,表明人类不得不再次以狩猎为主。
农业起源于中东。但是,这些植物品种,起源在哪里?
病态的农业起源
俄罗斯最著名的植物学家和遗传学家尼古拉·瓦维洛夫(Nikolai Vavilov ,1887-1943)游历欧洲等地学习之后,在俄国组织了探险考察队,搜集了世界上几乎每一个角落的植物种子,存放在彼得堡。这个资源库至今依然是最有价值的植物资源库之一。
瓦维洛夫详细分析了世界上主要粮食品种的起源。最大的一个起源地在中东的新月沃土地区,其他几个起源在伊朗——印度——中国——东南亚、中美洲——南美洲安第斯山脉等地。这些起源地有一点相同,即全部起源于山区。山区是生物学多样性的避难所。气候导致的各种灾难和干旱,造成其他地方的植物灭绝时,山区的植物却幸存下来。人类从山里挑选出可以作为食物的植物品种,这些粮食品种被世界各地的人类带下山的时间基本上差不多。
中美洲的玉米、土豆、豆类、辣椒、巧克力、香草、菠萝、南瓜都起源于墨西哥南部的瓦哈卡(Oaxaca)附近。瓦哈卡的地形崎岖不平,形成很多山间的大小盆地,这里的文明出现于一万年前,起源于这里的最重要的粮食作物是玉米。玉米迅速向四处扩散,进入北美和南美,正如小麦从新月沃土的扩散一样。
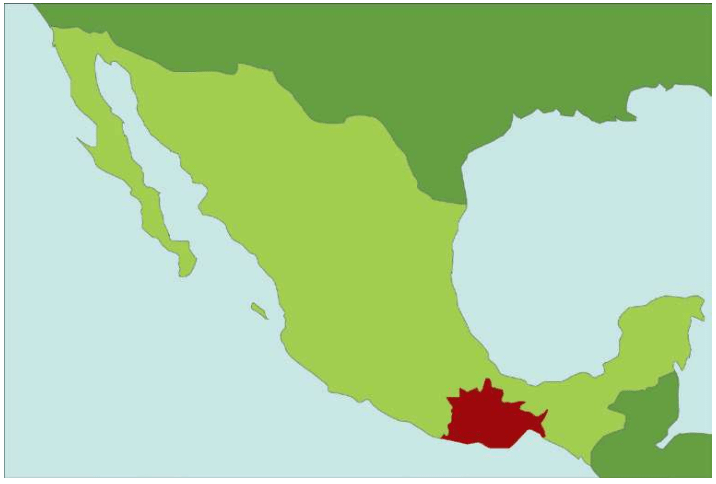
在北美人的骨骼遗骸里,突然出现了新的“碳信息”。
植物的碳分子结构并不相同。2.5亿年前,出现C3植物,只有3个碳原子。这类植物用大气中的二氧化碳生产3个碳原子的植物分子,利用光合作用储蓄能量。现在世界上大约95%的植物属于C3植物,狩猎采集群体食用的大部分植物也都是C3植物。6 500万年前,植物又进化了,出现C4植物,拥有4个碳原子的分子,包括大部分热带草本植物,例如玉米、谷子、甘蔗等。
大部分碳分子有6个质子和6个中子,6+6=12个原子,亦即碳12。但是少数碳分子拥有7-8个中子,形成碳13和碳14同位素。植物死亡后,残留的碳分子会逐步丢弃多余原子,回到12个原子的正常状态。碳14被丢弃的速率不变,检测碳14的残留量可以推算时间。同时,碳13继续维持不变。可以通过检测碳14的存在比例了解历史。考古学家就是用这种办法,检测出玉米的信息出现在北美土著的遗骸里——他们吃的C4食物(例如玉米)越多,碳14的比例越低。
根据骨骼的年代和碳14的比例高低等数据,科学家查清了玉米传播的大致途径和时间:北美的农业,出现时间还不到2 000年。随后,“玉米杀手”迅速传播。
水稻则起源于中国南部。水稻的野生祖先野生稻(Oryza rufipogon),至今仍然生长在印度和巴基斯坦,在中国和北美也有野生稻。
经总部设在美国的史密森尼学会(Smithsonian Institution)仔细研究发现,在1.3万年前,中国长江中游出现过水稻,但是在北半球突然变冷的“新仙女木”时期却消失了,留下了植物化石。1.1万年前,水稻的植物化石又出现了。所有的粮食作物的起源必经的最后一个步骤就是驯化。

威廉·布莱船长(William Bligh,1754-1817)的故事《叛舰喋血记》(The Mutiny of the Bounty)曾5次被搬上银幕。1789年,他率领的“邦蒂号”(Bounty)经过6个月航行抵达塔希提。他一路上都严苛地虐待水手,抵达塔希提后,他强令水手不许寻找当地女人以免传染性病。
威廉·布莱当时在塔希提滞留的原因是面包树(breadfruit)。这种植物美味可口,富含卡路里、易于生长,但是却没有种子,只能在树枝上砍出缺口,缺口长出根系后,砍下这段树枝移植到其他地方,最后成为一棵独立的树。
布莱带着两个植物学家耐心地培育尽可能多的小树,准备用船带到西印度群岛种植。这是一个漫长而枯燥的过程,饱受虐待的大部分船员叛变驾船离开,只有少数忠于他的船员留在塔希提。
挪威的海上丰收公司(Marine Harvest)的三文鱼人工培育过程也采取这种“闭合循环”的反复重复。繁育过程中,没有外来的植物和动物的介入,原来的物种本身不断反复循环,最后出现很多不同的变种。
野生鳕鱼4-6年才能长大成熟,海上丰收公司重复繁育,找到了两年成熟的一个新品种。这家挪威公司的大部分预算都投入在重复繁育。为了培育新的比目鱼,他们甚至建立了很多海水养殖的“比目鱼大厦”,让更多比目鱼住进楼房,从而容纳更多的重复繁育。
现在看看三大谷物:小麦、水稻、玉米。
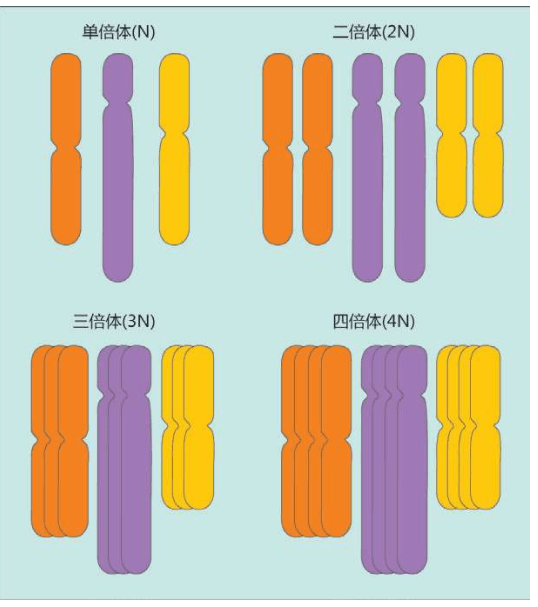
这三种谷物提供了世界人口的大部分卡路里,但是,现在它们都是同源多倍体(polyploid)——它们已经被人类繁育了很多代。换句话说,小麦、水稻、玉米的基因组重复了很多次,染色体变成原来的很多倍,它们成了转基因植物。
现代的转基因范围更广泛了,已经发展成移植抗病、抗虫的基因等。
正常情况下,一个基因组里的染色体的数量大约上百万年才会增长一倍。人类的驯化使得小麦、水稻、玉米的染色体数量翻了很多倍,这是相当于几百万年才能有的进化。
复制录像次数越多,图像越模糊,复印资料也会发生错误或遗漏某些页——复制基因组当然也会发生很多错误。基因组染色体不匹配的植物无法产生后代,复印时必须保留一份“原件”,以备再次复印——第二年作为种子播种,这是多倍体谷物的缺陷。所以农民必须购买种子,否则,产量会一代又一代地自动退化回去,这是一种源自基因的力量。
人类这种反复的重复繁育,存在致命的风险。
第一个指出这种风险的学者是日本裔美国生物学家大野乾(Susumu Ohno,1928-2000),他在1970年所著的《基因重复的进化》(Evolution by GeneDuplication)一书中提出:重复基因时,随心所欲地草率选择,会导致“快速进化”的变异,必须保留备份。他创造出“垃圾DNA”(junk DNA)一词,用以描述基因组里的很多功能不详的DNA。这种垃圾是重复基因的必然宿命,也许毫无意义,也许后果致命。
正常发挥功能的基因维持生物体“继续活着”,无法预料的突变则可能夺去生物组织的生命。所有癌症都是不断重复自己的DNA,最后发展成为同源多倍体的。
这种危险的同源多倍体的小麦、水稻、玉米呈现出极高的突变率:它们的DNA始终处于不稳定的、重复的、分子湮灭的状态。
分子湮灭时会产生基因换位现象(transposable elements),成为DNA上寄生现象的起因。病毒可以嵌入基因,尤其是活跃的逆转录病毒(retroviruse)。艾滋病毒(HIV)就属于逆转录病毒家族。1950年,美国女遗传学家芭芭拉·麦克林托克(Barbara McClintock,1902-1992)在研究玉米遗传时发现了这一现象。起初遗传学界表示怀疑,后来证明她是正确的。
现在的玉米分为很多类型,包括双倍体、四倍体、多倍体等。玉米的最重要的3个基因分别决定玉米棒的数量、玉米颗粒的层次数量和玉米的糖分。经过仅仅4 400年的人工选择,尤其是最近2 000年的培育,玉米已经出现巨大的变异。
在人类居住了4 000年的叙利亚境内的阿布·胡列伊拉遗址(Tell Abu Hureyra)中,出土了1.1万——0.75万年前的人类留下的150类500多种食用植物种子,研究这个遗址的学者们认为起源的定义有几种,人类刻意选择物种也算是一种“起源”。
如果某些物种没有产生人们期望的后代,人类就从野外再找其他物种。这些中东的早期农民,最后在“150类500多种食用植物种子”中留下了仅仅8类农作物,其中就包括小麦。
开展农业是人类的一个历史性决定。向大自然开战,与几百万年的进化史告别,人类与人类本身也决裂了——没有任何计划和目标的远征开始了。在没有地图和导航的黑暗中,人类跌跌撞撞地开始前行。
农业文化,带来两个重要的新事物。
第一,农业带来了人口数量的第二次1 000倍数量级增长。
第二,农业催生了政府。
狩猎采集者之间是基本平等的。农业却使人类无法离开耕地和水源,定居带来各种建筑和不动产,大量人口群集造成不同的社会阶层和不平等。世界上的所有人类群体都自发诞生了宗教。为了合理分配水源和土地、联合开凿引水渠、共同建设神庙……人类突然发现必须有一个新的东西——政府。
是农业文明最终带来了政府。
政府开始组织更大规模的农业生产,从而产生出更多的人口。人口压力再迫使人们继续迁移,寻找更多的土地和水源。政府的诞生,原本为了协调农业生产,但很快转变成为战争组织,以夺取新的土地。最后,政府演变成帝国,如埃及、亚述、波斯、希腊、罗马、印度孔雀王朝、中国汉朝、高棉帝国、大津巴布韦等。
本质上,政府的诞生源自生育的力量。为了争夺土地资源,政府组织的战争越来越多,规模越来越大。随着战争日益频繁和残酷,社会主导地位向男性倾斜,战争英雄成为被崇拜对象或政府首领,而发明农业的妇女却沦为农业社会的下层。(如尼罗河两岸原本有40多个政府(Nome,诺姆),经过几千年的残酷战争后兼并为两个政府:上埃及和下埃及。5 100年前,上埃及的法老那尔迈首次统一两个埃及)
农业使我们病了
世界自然遗产大烟山国家公园(Great Smoky Mountains National Park)是美国旅游人数最多的国家公园之一,每年有900万——950万游人。位于田纳西州东部的大型游乐场多莱坞(Dollywood)的游客每年超过200万人。如果我们去大烟山和多莱坞旅游,就会发现几乎处处都是肥胖者。
虽然大烟山的游客来自全美和世界各地,但是大部分还是来自附近各州。这些州的肥胖比例超过20%,密西西比州的比例更是超过三分之一。肥胖比例最高的州都是家庭平均收入最低的州。但是,美国的东海岸、西海岸和欧洲,也是肥胖者越来越多,很多场所的座椅尺寸已经无法容纳越来越庞大的人体。
肥胖已经不是一种现象,而是一种疾病,一种流行病。
1991年,美国没有任何一个州的肥胖人口超过20%。仅仅20多年间发生的变化无法用基因变化来解释。现在85%以上的美国人认为,肥胖是一种病。美国疾病控制预防中心(Centers for Disease Control and Prevention)和世界卫生组织的调查确认,肥胖是仅次于吸烟的第二大流行病,并将在10年内成为世界第一大流行病。(现代人的食物远远超过了实际需求。线粒体以氧气为原料,每天制造的ATP能量的重量占人体体重的一半,为人类制造能量的效率为20万倍)
肥胖是很多疾病的基础。
我们的丰富食物,既滋养了我们,也在杀死我们。
欧洲的肥胖病也在流行,发病率最高的地区也是经济最不发达的地区。这和美国的统计完全一致。
发展中国家的肥胖病也在流行。过去的解释认为,收入低的地区,受教育程度低,进入社会后的收入也低,成为恶性循环。因为运动缺乏,食物过量,才导致肥胖。现在,这些说法已经不足以解释全球性的肥胖病,因为收入中等、人口最多的印度和中国也在受肥胖病及其带来的诸多恶果困扰。
政策制定者和公共卫生专家已经认识到,除了糖尿病,肥胖还会引发一系列严重的慢性疾病。印度金奈的医学家们发现,人类很难改变固有的生活方式,贪吃和不爱运动既是强力的毒品,也是人的天性。
美国遗传学家詹姆斯·尼尔(James Neel,1915-2000)在对广岛和长崎遭遇原子弹辐射的群体研究中曾经惊讶地发现,基因突变率没有增加,也就是说,甚至强烈的原子辐射也无法撼动DNA。然后,尼尔着迷于巴西——委内瑞拉边境的原始土著——亚马孙流域的亚诺玛尼人(Yanomami,约两万人的部族)的遗传研究。这里原来与世隔绝,人们生活在天然状态,但随着转入农业的进化压力,基因却很快发生了显著变化。
尼尔猜测,在狩猎采集群体里,原先几乎没有糖尿病,所以这种疾病可能是突然大量供给卡路里的一种生理反应。卡路里的低摄入是狩猎采集群体在长期进化中形成的一种维持正常生理机能的能力,这种能力与现代的丰富饮食发生了严重冲突。他称其为“节俭基因型”(thrifty genotype)。
这个理论,随着糖尿病日益普遍得到认可。显然,这又是一个自然选择的行为。
糖尿病分为两种。Ⅰ型糖尿病可以在儿童时期发病,病因是DNA的遗传因素与环境的综合。Ⅱ型糖尿病在成人时期出现(也有少年时期的发病者),部分源自遗传,部分源自日常饮食,80%以上的Ⅱ型糖尿病患者都超重或肥胖。
这种“节俭基因型”副作用的最有趣的例子,是太平洋波利尼西亚诸岛屿的居民。萨摩亚人(Samoans)在3 000年前从亚洲南部迁徙到这些岛屿,他们有时必须承受几个星期营养不足的强大生理压力,幸存者属于能够减少卡路里消耗的群体。他们从事农业,获得丰富的卡路里,但是他们几乎天天四处活动消耗热量,防止了肥胖的流行。
但是,现代文明来到后,萨摩亚人不再乐于田间劳作和出海捕鱼,大部分时间坐着享受生活。现在,城镇的萨摩亚人中的肥胖者超过人口的三分之二,乡村中的肥胖者超过一半。所以萨摩亚人的糖尿病比例很高:男性25%,女性15%。
在美国西南部和墨西哥北部的美洲土著比马人(Pima Indians)中,生活方式与糖尿病的密切关系更令人恐怖。住在美国的比马人,物质丰富,高达40%的人口罹患糖尿病,这个比例是全世界最高的。但是,边境另一边的同一族群却较多地维持着传统生活方式,只有7%的糖尿病患者,虽然这个比例也不低,但远远低于美国比马人。
现在人人皆知“过度饮食和缺乏运动导致肥胖”。我们的先祖在狩猎时必须节省每一个单位的卡路里,因为每多消耗一个单位的卡路里,就必须多猎取一个单位的卡路里进行补偿。锻炼身体对他们来说是荒唐可笑的,他们本来就在奔走狩猎或四处采集种子和水果。这种几百万年的进化,产生了一套极其高效率的能量存贮和使用能力,早已深深埋藏在我们的基因里。
所以,糖尿病的威胁,不是来自外部,而是来自内部。
对肥胖——糖尿病的理解,涉及基因的进化及文明的发展历程。肥胖——糖尿病已经引发了无数后续的疾病。美国政府的统计发现,肥胖引起的各种疾病在人均医疗支出中的比例已经稳居第一位,约占四分之一。
第三次疾病浪潮
2003年2月,一位中国医生到香港出席侄子的婚礼,他无意中散播了一场蔓延全球的传染病。他当时感觉自己病了,以为得了感冒。这种感冒可以通过空气快速传染,最后出现肺炎,死亡率约10%。当时,这种疾病迅速扩散到五大洲,世界旅游行业下跌了9%。这种疾病源自冠状病毒(coronavirus)的一种,从此,一种新的传染病SARS(Severe acute respiratory syndrome,严重急性呼吸系统综合征)诞生了。
SARS的特点是传播速率惊人。这种病可能是中国南部的鸡病毒或猪病毒的突然变异。宿主成为人类后,通过饲养场的雇员和这位医生,一个月里就传播到加拿大、瑞士和南非。
SARS的死亡率不高。非洲的埃博拉(Ebola)、拉沙热(Lassa fever)和马尔堡病毒(Marburg virus)的死亡率极高,但是,这几种疾病仅仅在相对封闭的群体里突然暴发,没有快速传播造成严重威胁。H5N1禽流感(H5N1 avian flu)的死亡率可以高达50%,顾名思义,这种病毒来自家禽。1918-1919年暴发的流感,在欧洲等地造成2 000多万人死亡。
这些传染病全部起源于人类饲养的动物,并非“新威胁”。
美国历史学家威廉·麦克尼尔(William McNeill,1917-)在他1976年出版的《瘟疫与人类》(Plagues and Peoples)一书中,阐述了疾病与人类历史的关系,他发现传染病是很多重大历史事件的催化剂。
例如,14世纪欧洲流行的黑死病迫使蒙古帝国的军队逃离,而西班牙对美洲各个帝国的成功征服,主要是欧洲带来的疾病造成了大部分美洲土著死亡。
麦克尼尔发现很多疾病起源于新石器时期:农业人口集中在较小的空间,导致疾病的发生和流行。大约一万年前,中东开始驯化绵羊、山羊、牛和猪。大约8 000年前,东南亚开始驯化鸡——人类第一次与其他动物生活在一个社区里。
麦克尼尔写道:“人类与他们驯养的动物分享各种疾病,26种疾病来自鸡,42种疾病来自猪。”
人类与牛分享的天花曾经杀死了无数人类,直到人类用种牛痘的办法遏制了天花。从动物传染给人类的其他著名疾病还有麻疹、肺结核和流感、黑死病。考古发现,人类过去从未罹患这些疾病,这些疾病通称“动物疾病”(zoonotic diseases,希腊语zoon的意思是疾病)。
在人类几百万年的进化中,体内长期存在多种细菌和病毒,但是它们与人类是互利互惠、寄生共存的关系。比如消化系统里的多种菌类,它们不会暴虐地杀死自己的宿主,断绝自己的生存资源。
考古证据发现,旧石器时代人类死亡的主要原因不是疾病,而是外伤和外伤引起的感染。外伤主要是狩猎造成的。进入农业社会之后,外伤导致的死亡下降,疾病导致的死亡上升,后来慢性非传染病开始大量增加,这三大死亡原因形成三次浪潮。
麦克尼尔的证据,引来很多类似的研究著述的出版。
从这三次浪潮的三条曲线可见:
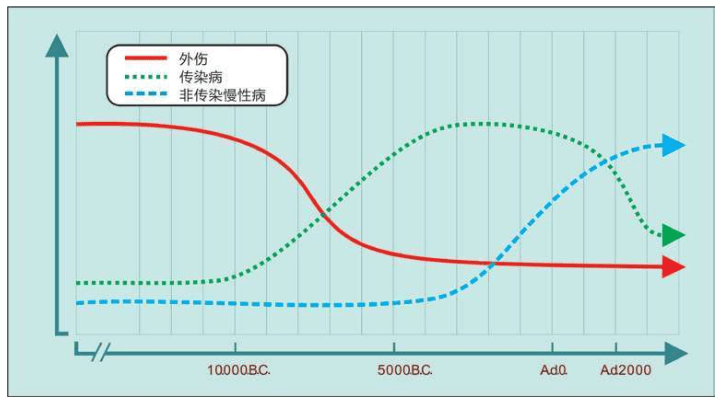
1.人类从旧石器时代到新石器时代,外伤是第一位的死亡原因。
2.定居和驯化动物以后,传染病成为主要原因,直至20世纪出现抗生素。
3.最后一条曲线发展若干世纪后,在20世纪中期上升——非传染慢性病如癌症、糖尿病、高血压、心脏病等成为人类的最大威胁。
造成这些现象的另外一个原因是医学的发展延长了人类的寿命,很多慢性病往往与年龄关联。但是,最严重的问题是三次浪潮中最新的一次巨浪——非传染性慢性病,这些疾病全部无药可医,换句话说,必须终生服药。
非传染性慢性病的起源是碳水化合物——糖,糖给人类带来疾病,其证据最早是从印度河谷出土的蛀牙化石发现的。这个证据,首先刊登在《自然》杂志上。
梅赫尔格尔(Mehrgarh)距离阿拉伯半岛3 000多千米,位于一个陡峭的山峰下的坡地。这里发现的最大一个村庄的面积达到2平方千米。这是世界最古老的新石器时代定居点之一,也是亚洲南部最老的人类定居点,5 000年前印度河谷文明的发源地。从梅赫尔格尔向东,还有多处古代村镇遗迹,其中摩亨佐·达罗(Mohenjo Daro,乌尔都语,意思是死城)是一座古城,被列入联合国世界文化遗产。
人类在梅赫尔格尔居住了大约4500年,这里发现了人类驯化小麦、大麦、牛、羊、山羊等的遗迹,以及石器、铜器、铁器等。9000年前定居在梅赫尔格尔的人类住在泥砖房子里,制作陶器,用海产与800千米外的帕米尔高原上居住的人类进行贸易。
在这里最令人惊讶的发现之一是古代牙科手术的证据,它来自9000-7500年的土层。那个时期的牙钻应该是石器,这是世界最早的活人牙科手术。
蛀牙,在旧石器时代的狩猎采集群体里几乎没有发现,在新石器时代才开始显著增加。它正是人类食用碳水化合物——多糖类食物的后果之一。为了减轻蛀牙的痛苦,史前的梅赫尔格尔的人类用石器做成钻头,磨掉被侵蚀的蛀牙部分。否则,严重的蛀牙会导致整个口腔的溃烂。北美土著的情况与此类似。研究发现,在狩猎采集生活方式中,发生蛀牙的比例不到5%。而进入农业时期以后,蛀牙增加到25%以上。
在农业时代,日常饮食里突然增加了大量的碳水化合物,这种食品必须经过去壳、研磨、发酵、烹饪等处理。在这些处理过程中,植物原有的很多营养丢失了,碳水化合物仅仅是糖。但是狩猎采集群体直接吃草类,不仅保持原有的营养,还起到了清洁牙齿的作用。
现在流行的阿特金斯饮食法(Atkin s diet)、旧石器饮食法(Paleo Diet)和其他版本的各种新饮食方式,都是回归狩猎采集时代我们先祖的饮食方式——新石器时代以前、农业出现以前的生活方式。(旧石器饮食法(Paleo diet):又称洞穴人饮食法(Caveman diet)、狩猎采集者饮食法(Hunter-gatherer diet)。这类饮食排斥谷物、豆类、乳制品等,推崇水产、食草类动物的肉类、蔬菜等)
美国医生罗伯特·阿特金斯(Robert Atkins,1930-2003)为了解决自己的超重问题,研究出不吃碳水化合物和含糖食品,只吃肉类、蔬菜的减肥饮食。1972年他出版了《阿特金斯医生的饮食革命》(Dr. Atkins’ Diet Revolution),此后又出版十余本同样理念的书,号召人类回归自然,避免非传染慢性病。这套办法风靡至今。(阿特金斯饮食的基本理念:可以摄入和消化脂肪,但是排斥碳水化合物和含糖食品,以免多余的碳水化合物变成脂肪存贮起来。阿特金斯饮食可能过分强调以蛋白质取代碳水化合物)
新石器时代的纳图夫人的食物中已经包含一定数量的碳水化合物。但是,进入农业时代之后,突然变成以碳水化合物为主。现代人类的食物主要是碳水化合物和脂肪,这确实完全背离了人科生物几百万年的进化史。
当然,看起来似乎淀粉比糖好。实际上,淀粉也是糖。
人类食用糖的历史已有几千年,蔗糖的规模生产起始于产业革命。欧洲原来用蜂蜜作为甜味调料,蜂蜜营养全面,包含多种维生素和矿物质,但是产量少、价格高,无法作为卡路里的长期来源。于是,产量大、价格低的淀粉成为卡路里的来源。
现代饮食中,排名第一位的罪犯是糖。人类的基因因为无法处理过量的糖(碳水化合物),从而导致糖尿病。另一个重要罪犯是添加剂。2002年,埃里克·施洛瑟(Eric Schlosser,1959-)出版了大型调查报告《快餐民族:所有美国人食物的黑暗面》(Fast Food Nation: The Dark Side of the AllAmerican Meal)。这本书列举了很多数据,例如,麦当劳草莓奶昔由60多种添加剂构成,唯独没有任何草莓成分,含糖很多。又如,番茄酱(ketchup)的三分之一是糖。这本调查报告引起巨大轰动,美国涌现大量类似书籍,出现多部电影,批判反思现代饮食文化。
人类在几百万年的进化中,形成了敏感的味觉和摄取偏好。对于苦味,人类本能地警惕,因为这些植物可能有毒。对于甜味,人类本能地感觉安全,因为它属于成熟的水果的味道。这些味觉和摄取偏好最后导致人类走向过量的糖——碳水化合物。偏好甜味正是人类进化史中的“阿喀琉斯之踵”(Achilles’ heel)。
19世纪的产业革命是人类的第二次产业革命,新石器晚期的农业革命是第一次产业革命——由政府组织实施的获得食物的产业化革命。梅赫尔格尔发现的人类蛀牙,只是农业革命的恶果之一。
过去的一个多世纪里,美国食物中的糖类大大增加,汽车取代了走路……美国各个年龄组的肥胖人数都增加了大约10倍。现在,电脑、手机、互联网和游戏机使得人类的体力活动和身体锻炼更少,很难预料未来将是什么样子。有的媒体估计,2000年出生的孩子,在2050年时大约三分之一会成为肥胖者。
所有这些疾病的根,都源自农业文化——我们要为对抗大自然付出代价。
糖尿病、高血脂、高血压、心脏病等非传染慢性病,要么无药可治,要么必须终生服药。这已经完全不是传统意义上的“疾病”的概念,这些症状,也源自农业文化——我们要为对抗人类的基因付出代价。
在发展中国家,情况也不乐观。中国的各种“吃”出来的疾病也在不断增加,以肝脏为例,城市中的肥胖和脂肪肝人群非常普遍,中国特有的地方病乙肝的数量已经超过三亿人口。这一切都发生在最近几十年里。
21世纪是生物世纪,但是不应该成为医药世纪。
2011年,世界的新药品中74%已经是生物药品。生物药品迅速超过了化学药品,这些药品中不仅有治疗身体疾病的药品,还有治疗精神疾病的药品。
难道我们六万年前走出非洲,就是为了今天吃药?
农业使我们疯了
人类走出非洲的旅程中,农业不仅引发身体疾病,还引发了精神疾病。
现代人头脑聪明,能够制作各种石器和武器,通过语言沟通和社会组织进行有效的狩猎和采集。现代人在非洲发展出这些文化,但是,他们为什么离开非洲?以前又为什么留在非洲呢?
对非洲各地的湖泊沉积物的数据分析发现,七万年前,最后一次冰河期,气候日益趋向寒冷,非洲也越来越干旱,草原减少;8 000年以前,非洲越来越干旱,撒哈拉沙漠开始扩张。但是干旱可能不是唯一的原因,人类曾经遭遇过一次巨大的天灾。
印度尼西亚的苏门答腊北部的多峇湖(Lake Toba),长度100千米,宽度30千米。这是一座休眠火山口,曾经发生3次大爆发。第三次喷发在七万多年前,是200万年里地球上最大的一次火山爆发,在大气层形成大量尘埃,遮挡了阳光,使气温急剧下降。火山灰散布到周围各地,造成全球性的巨大生态灾难。直到今天,在印度中部一些地区还覆盖着厚达6米的火山灰。
当时已经进入冰河期。火山灰遮蔽阳光,导致全球出现6-10年“火山冬季”(Volcanic winter)和大约1 000年的极度严寒,非洲地区的气温进一步下降。斯坦福大学的研究发现,当时非洲的人类数量减少到几千人,濒临灭绝。而其他地区的智人亚种受到了更大的打击。欧洲的尼人完全灭亡,西亚的尼人只留下高加索山谷中数百人。东亚的丹尼索人也可能只剩下横断山区数百人。多峇火山爆发为现代人走出非洲扫清了障碍。我们仍然不清楚当时发生了一些什么故事。但是,再次发现人类时,他们的石器和艺术已经大不相同,人类文化发展了——这种进步明显受到了强大的进化压力。很多学者猜测,这次多峇火山爆发可能是人类进步和走出非洲的一个非常重要的刺激因素。
这个理论被称为“多峇巨变理论”(Toba catastrophe theory),又称多峇突变理论:巨大的自然灾难带来瓶颈效应,迫使人类发生了突变。
进入工业化之后,人类成为马歇尔·萨林斯(Marshall Sahlins)激烈批评的“经济人种”(Homo economicus,或经济动物):每天受时间的严格约束,无数人天天重复做同一件事情,探险精神和创新精神成为少数人的专利。
进入现在的后工业化时代,人类“进化”为不同的 “专业人种”:
在微粒子物理论坛上,一个民事诉讼律师根本搞不懂物理学家们在说什么。
在文学理论学术会上,一个化学家也是多余的听众。
每一个人都不会修理自己的汽车和家用电器。
每一个人都在过着食品和娱乐过剩的生活,但还是像机器一样在拼命。
……
我们已经找不到自我,找不到生活。
1997年,托比·莱斯特(Toby Lester,1964-)发表的文章说:“我们是机器噪声包围的第一代人类。”无论在工厂还是办公室,各类机器设备比比皆是,家里的家用电器遍布每个角落,互联网和手机须臾不可分离……听得见的噪声、听不见的电子噪声、无数的社会联系,正在悄悄地损害着我们的免疫系统。
人类能够走遍世界,首先是因为具备了强大的身体和精神力量。所谓身体的强大,并非肌肉的强劲,而是免疫系统的发达。在一个细菌病毒无所不在的世界,任何幸存的物种都必须拥有强大的免疫系统。所有人属物种都灭绝了,因为人类的免疫系统最强大,所以只有现代人幸存下来。但是,现在越来越多的癌症正是免疫系统无法控制细胞基因突变的一个最好例证。
狩猎采集时代的田园牧歌,不可能时光倒流。(北美土著有一句古老格言:善等地球。它不是你父母给你的,它是你的孩子们借给你的。Treat the earth well. It was not given to you by your parents, but is loaned to you by your children.)
每一个民族,都曾经回忆和怀念远古的传说,那是人类的黄金时代。探索和反思人类六万年的旅程,只能得出一个结论或者疑问:我们是不是疯了?
著名的“邓巴150”是精神疾病起源的另一个证明。
考古学家曾经研究分析过尼罗河沿岸的历史。干旱驱赶着日益扩大的撒哈拉地区的无数部落氏族,拥挤到狭窄的尼罗河两岸。资源的争夺与战争不可避免,在这些资源争夺战中,有的埃及部落死亡率超过40%。但是,狩猎采集时代,人类还有更多的空间,最好的出路是走开,离开尼罗河,走向欧亚大陆。实际上,《圣经》也是描述了犹太人走开的一个例子。
猴子、猿人和猩猩也会发生资源冲突,它们的最终选择也是走开。
进化心理学家罗宾·邓巴(Robin Dunbar,1947-)对复杂社会架构的大量群体中,如何缓解压力感到好奇,于是研究了各种猴子、猩猩等灵长目动物的平均群体数量。他发现,平均群体数量与大脑皮层有关,大脑越大的灵长目动物,迁移和生活的群体数量越大,因为神经元的增加,扩大了可以联系的个体数量,即社交联系的范围。大部分物种的群体数量在5-50只个体。他推算出人类大脑与相应的联系个体的平均数量为148个人,这个数据后来被称为罗宾·邓巴150(Robin Dunbar 150)。
罗宾·邓巴解释:这个数值是人类理想的社交联系人数,虽然有人最多可以记住2 000个人,但是保持有意义的社会关系的人数是150。这个数字正是1990年代发明E-mail之前,人们在圣诞节邮寄贺卡的平均人数。
这是生物学决定的一个“硬链接”数值。如果超出这个数值,只有两种可能性:要么分离,要么调整和改善政府、宗教、法律、警察等社会组织的相应结构以维持这个数值。
旧石器时代的先祖可以选择分离,我们只能选择后者——调整和改善。
无论怎样调整和改善,现代社会的社交人数,还是往往远超150人,由此也带来心理负担和精神压力。
农业发展、产业革命、互联网时代使得人类社会变得如此复杂,我们陷入了一种既不可能“非人化”,也不可能违背自然规律的两难境地。于是,我们不愿观看身边的人海,宁愿在公共场合埋头玩手机;我们对各种社交聚会犹豫不决,又不得不前去社交和应酬……事实上,我们的大脑根本处理不了这么多的“社会关系”,也没有时间安排这些“社交应酬”。
这种大背景,正是精神疾病日益增多的另一个原因。国际卫生组织已经将这类疾病列为2020年世界范围内第二大的致残和致死原因,在欧洲和北美,出现这类精神疾病症状的人口比例都已超过四分之一。
复杂的社会结构和社会关系,使得人类在心理、精神和神经方面的疾病也成为一张长长的清单。
我们是不是应该收敛疯狂?
这种趋势的连带后果是精神疾病药物的使用在不断增长,人们试图改善自己的精神状态,酒精已经不能帮助人们缓解和摆脱精神的烦恼和压力……
根据美国疾病控制中心的统计,抗抑郁药类(如Prozac和Paxil)是美国现在数量最多的处方药,超过治疗高血压、胆固醇和头痛药剂的总和。此外,美国大约10%的男孩在天天服用含有兴奋剂Ritalin的药物,以控制注意力不集中和失调多动症状……
虽然新的更有效的药物正在继续研发,但是无法解决农业文化和人口爆炸这两个孪生的病源,因为我们不可能战胜我们自己的基因。
这是人类历史上的第一次。
时光虽然不能倒流,但是我们可以回到狩猎采集时代,看看几百万年进化的人类,找出真正合适的社会交流范围,以保护我们的免疫系统和健康。
我们应该学习了解生物学和基因的新知识和新发现,我们应该参考农业出现之前的狩猎采集时代,再造新的文化,再造人类。
已经开始的溃败
农业扩张一万年后,人类第一次开始从土地上撤退了。
图瓦卢(Tuvalu)是一个太平洋岛国,由9个珊瑚礁环岛屿组成,面积约1.054万平方千米,26万人口。这个岛国最高海拔仅5米,随着全球大气变暖,南北极冰盖融化以及全球冰川消退,欧美各国在图瓦卢安设的精确设备测量发现,太平洋海平面平均每年上涨0.9厘米,预计21世纪之内,也许50年内,图瓦卢将淹没在大海里。
随着海平面上升,这个岛国现在已经无法获得淡水,所有食品和饮水必须进口,导致各种包装垃圾遍布各个岛屿。图瓦卢的旅游业已经消失了。
在德国、法国、英国、美国、日本、澳大利亚等国的帮助下,图瓦卢一面接受国际援助,一面进行谈判和规划,他们只能放弃家园、举国迁移。
这是人类历史上的第一次国家整体溃退。可怕的是,这一现象,正在全球蔓延。
人类终于认识到索取地球无止境的农业文化的重大内在缺陷,觉悟到农业文化贪婪的恶果。人类开始退耕还林,退草还牧,控制人口。
20世纪,人类最重要的发明之一是退休,退休之后由国家福利体系养老。
退休制度的重大成就之一是遏制了世界人口的继续迅速增长,人类不必为了吃饭、为了养老而没有节制地大量繁衍下一代。
20世纪,发达国家人口保持稳定的事实,已经证明托马斯·马尔萨斯的政治经济学理论是正确的。他的理论深刻揭示了人类与大自然的关系。农业文化的本质是无止境地索取地球、繁育人口,形成恶性循环直至地球资源枯竭。
我们必须改变农业文化以来的世界观。
所谓“开化的”农业始终是野蛮的。
最初的农业是“刀耕火种”:破坏和利用一块土地,然后迁移到另一块土地。农民播下种子的目的,是为了收获和养活家人,土地只与生存有关。土地不足的时候,为了争夺土地资源,就会发动有组织的战争,互相屠杀。进入“文明的”工业时代,人类“进步”了,开始更加努力地增加产量——向地球索取。这种循环没有止境,直到再也没有地方可以开发,直到世界再也没有新的资源,人类再也无处可去。(我们人类根本不需要那么多粮食、肉类、蔬菜和水果)
农业文化认为,向地球索取可以无穷无尽,尤其最近几个世纪的无限制扩张和掠夺几乎达到疯狂。可是,土地终有尽头,地球终有尽头。
农业文化发展进步的陈旧模式面临资源枯竭的致命挑战,继续维持已不可能。虽然我们无法回到农业以前的时代,但是狩猎采集时代的人类文化值得我们反思和借鉴。
人口与人,完全是两个不同的概念。托马斯·马尔萨斯(Thomas Robert Malthus,1766-1834)说:“人口的力量无限大于索取地球而求生存的人的力量。”
人是猎人,几百万年的猎人。
六万年前,我们离开非洲。
一万年前,潘多拉的盒子打开了。
我们的进步和错误,都发生在一万年之内。基因研究告诉我们,我们是不可分割的远亲,人类也只有一个地球家园。
我们不可能回到狩猎采集时代。在陆地资源已经所剩无几,当我们继续带着农业文化的世界观,准备走向海洋继续索取,继续探索基因技术以求新的发展之前,是否应该首先勇敢地再次审判自己:我们做错了什么?
仅仅20多年,生物科学和基因技术揭示出一系列事实,使得人类陷入全面的反思和探索。
第八章 必须向新理论开放
人类科学研究的历史证明,我们必须向新的理论开放,因为新的理论可能被新的事实证明。在研究人类的遗传、人类的起源、人类的旅程、生命的起源的过程中,一百多年来涌现了无数不屈不挠的先驱者,他们在一团谜雾中摸索,他们一次又一次否定自己、发现真相,他们带领我们找到了人类的先祖,了解了人类波澜壮阔的六万年旅程。
这一章,我们主要介绍其中最重要的几个人物,几个诺贝尔奖获得者的故事,以及最新出现的新的生命理论。
这些内容,都与本书的主题、人类六万年的旅程息息相关。
基因的先驱与DNA的先驱
中世纪,“先成者说”(preformationist theory)认为,人类本来就在卵子里。如果“先成者”存在于母体里,那么,母亲的母亲的母亲的……母亲的体内,就应该有一个孩子的胚胎。这个理论有些像俄罗斯的套娃,它的真实含义是人类是在不断的退化过程中,所以这个理论被抛弃了。
精源论(spermists theory)和卵源说(ovism)更加糟糕。
按照《圣经》的创世逻辑和理论,上帝应该在创世的第六天,把所有人类的精子都塞进了亚当的身体里,包括亚当的孩子的孩子的孩子的孩子……的精子。这显然是不可能的。同样道理,夏娃被塞进无数世代的无数卵子当然更是不可能的。
科学家们相信,是某种化学物质参与了所有的遗传和进化。
基因与DNA,现在似乎已经成为同义词,其实并不一样。DNA是化学名词,基因是遗传学概念。
DNA——基因构成的大型分子结构叫作染色体,DNA结构蕴藏着基因的信息。
19世纪的中期,基因和DNA几乎被同时发现。两个伟大的先驱,基因的发现者格雷戈尔·孟德尔(Gregor Mendel)和DNA的发现者弗里德里希·米歇尔(Friedrich Miescher)都默默无闻地死去了。
1884年1月,孟德尔去世的那个冬天非常寒冷。修道院里修士们腾空了孟德尔的办公室,无情地烧毁了他的所有文件和院子里的实验设施。当时谁也不承认孟德尔的基因研究,没有任何亲友认领孟德尔的遗物,虽然后来作为遗传学之父的孟德尔成为这座修道院的无价之宝。
1884年,同一个寒冷的冬天,瑞士的米歇尔正在他的修道院地下室里进行三文鱼的实验。他的面前,放着他多年来从三文鱼的精液里提炼出来的一大堆黏糊糊的东西。他沉迷于这些实验,当年他的朋友们把他强行从这个地下实验室拖出来、前去参加他自己的婚礼。正是从这些黏糊糊的东西里,米歇尔提炼出了DNA。
孟德尔和米歇尔所在的修道院,距离大约600千米。但是,几乎整整一个世纪,没有任何人把基因和DNA联系在一起。
米歇尔的父亲是一位著名妇产科医生,因为米歇尔的耳朵不好,人们建议这个孩子从事化学研究。1868年,米歇尔进入生物化学家费利克斯·霍普-赛勒(Felix Hoppe-Seyler)的实验室。现代人无法想象,这个实验室竟然在一座古城堡地下室的王室洗衣房和厨房里。米歇尔做事非常专心致志,他取出德国伤兵伤口里的脓,在其白细胞的细胞核里,发现了一种化学物质。他用猪胃里提炼出来的酸溶解了细胞膜,分离出灰色的糊状物。但是他无法分辨这到底是什么东西?这种物质不是蛋白,因为和蛋白完全不同,这种物质在盐水、醋酸、稀盐酸里都无法溶解。他把这种物质命名为核酸(nuclein),后来学术界称其为DNA。
1869年,霍普-赛勒认为米歇尔的实验搞错了,要求他一步一步地再次重复这项实验,否则不允许他发表这项实验的结果。
1871年,又经过两年实验,米歇尔发表了他的论文。这篇论文,使米歇尔成为DNA的发现者。
1871年,同一年,达尔文的第二本巨著《人的由来》也发表了。
达尔文、孟德尔、米歇尔等遗传——基因——人类科学的先驱探索科学领域的时代,正是人类文明发生重大转折的时代,三场伟大的战争几乎同时在欧洲、亚洲、美洲进行:
1861-1865年:美国南北战争,双方动员军队超过300万。此战后美国成为真正统一的国家。
1868-1871年:日本明治维新,德川幕府退位,天皇从京都迁都东京,经两次全国性血腥内战“废藩置县”(300多藩主被废,改为70余县)后,日本成为统一国家。
1864-1871年:德国统一战争,俾斯麦三战三捷,先后击败丹麦、奥地利和法国,1871年,德国成为统一国家。
19世纪结束时,英美法德日五大强国完成工业化,科学技术高速发展。但是,生物——遗传——基因——DNA科学的道路仍然崎岖坎坷。
1869年,25岁的弗雷德里希·米歇尔(Friedrich-Miescher,1844-1895)在德国图宾根(Tubingen)一座古代城堡的地下室的简陋实验室里发现了核酸(Nucleic acid)。这种物质是DNA的构成单元。(同一时期,孟德尔发现了基因)
此后,米歇尔经历了一段不成功的教学生涯,因为“(学生)难以理解”和“性格焦躁”,米歇尔不得不离开教室,再次回到实验室。这一次,他的研究对象不再是伤兵的脓,而是三文鱼的精液,因为米歇尔发现三文鱼的精液几乎全部是DNA。他每年秋天和冬天都在莱茵河里捕捞三文鱼。他努力捍卫DNA的声誉,他的预算有限,他不被理解,他的努力也付诸东流——他无法回答这种神秘的DNA到底是什么?他也不知道这种物质如何影响了遗传?瑞士政府也不支持他。最后,米歇尔不得不全部放弃了DNA的研究。
当时,大部分人依然相信是蛋白质决定了遗传。
当时,大部分人认为DNA是不稳定的。所以,甚至米歇尔自己也开始研究蛋白的氨基酸如何影响遗传。最后,顽强的米歇尔发现,蛋白的氨基酸无法解释遗传,于是他再次转回DNA。不幸的是,他的身体越来越糟糕,1895年,米歇尔因为肺炎而撒手人寰。
孟德尔的遗传学研究更不顺利。他选择豌豆的原因之一是实验简单,昆虫和风都不会帮助豌豆授粉。孟德尔认认真真地记录每一天的温度和气压,以及各种豌豆实验数据,最后他的豌豆遗传实验几乎变成了一批统计学表格的综合。
1865年,孟德尔在一次会议上宣读了他的遗传学论文。史料记载,所有听众都认为这是一场数学计算演示,会议上没有出现讨论,甚至没有人提出任何问题。
1866年,孟德尔书面发表了他的结果。仍然是一片寂静,没有人回应。
1868年,孟德尔被选为修道院的院长之后,他一方面管理修道院,一方面仍然顽强地继续着他的实验。他过去每天抽20支雪茄,最后发展到最多一天抽120支雪茄。他晚年的一位来访者回忆,孟德尔曾经带着他参观修道院的花园和果树,但是看到豌豆的实验田的时候,孟德尔赶紧转移话题,甚至显得局促不安。来访者问他为什么种植豌豆?孟德尔回答说:“这只是一个小游戏,故事很长,长得一时难以讲清楚。”更加麻烦的是,孟德尔为了教会的利益与政府进行了长期的抗争,一位律师感叹地写道:“孟德尔的身边‘充满了敌人、叛徒和告密者’。”四面受敌,身体日渐衰弱的孟德尔担心自己被活着烧掉,所以坚持死后进行尸体解剖。
1884年,心脏和肾脏都失去功能的孟德尔最终死去了,修道院烧毁了孟德尔留下的一切,以维护修道院的名誉,豌豆实验也成为他的陪葬品。
孟德尔死后35年里,先后11位科学家(大部分不是农业科学家)分别表示,他们不认可孟德尔的遗传理论。这些科学家再次埋葬了孟德尔。
孟德尔曾经跟他的一位同事说过:“我的时代终究会来到。”1900年,3位生物学家几乎同时发现孟德尔是正确的。这3位科学家各自发表的论文印证了孟德尔的研究,他们不约而同地复活了这位捷克的修道士和他的基因理论。
但是,“基因”到底在哪里?仍然是一个谜。
胡戈·德弗里斯(Hugo de Vries,1848-1935),荷兰植物学家和生物学家,1901年发现突变(Mutation)并提倡突变理论。
埃里克·冯·切尔马克(Erich vonTschermak,1871-1962),奥地利农学家。
卡尔·埃里克·科伦斯(Carl ErichCorrens, 1864-1933),德国植物学家和遗传学家。
果蝇造就的一批诺贝尔奖
19世纪后期,达尔文的理论衰落了。生物学家承认演化发生了,但是他们贬低达尔文的自然选择机理,认为适者生存证据不足。人们认为适者生存的原理仅仅适用于不适者,演化是跳跃的或突然发生的,而不是达尔文所说的微小的演化。
19世纪后期,统计学进入生物学,人们发现个体特征(Trait)的变化往往呈现钟形曲线,例如,人群的个子都差不多高,特别高和特别低的人都是少数。显然,自然选择不能去除最高的人和最矮的人。
1900年之后,孟德尔主义(Mendelism)迅速流行,这种基因理论开始挑战达尔文主义(Darwinism)。人们认为这两种理论互不相容。
当时已经出现了染色体理论、突变理论、基因理论等,但每一种理论都不那么清晰和有条有理。科学家们互相争执,这些理论似乎互相重叠了。有人认为基因不在染色体里,有人认为一个染色体里只有一个基因……达尔文被忽略了。
达尔文及其支持者推翻了《圣经》,但是现在又出现了针对达尔文的战争。
1900年,一场不文明的内战打响了,这是孟德尔的遗传学针对达尔文的自然选择的战争。大部分生物学家认为,这场战争的结果将是一个理论灭绝另一个理论。三个复活了孟德尔的科学家之一,胡戈·德弗里斯(Hugo de Vries)首先发明了突变理论(mutation theory),他认为物种起源是某些罕见的突变引起的。
摩尔根(Thomas Hunt Morgan,1866-1945)原来是一个动物学家,研究胚胎。1900年,他听说了突变理论之后,照搬孟德尔的做法进行研究。他选择的不是豌豆,而是果蝇。当时果蝇移民到美国,香蕉也进口到美国。因为果蝇(drosophila,又称fruit flies)可以12天繁殖一代,所以成为一个很好的实验工具。
摩尔根在他的实验室里放了8个大柜子,几千只果蝇幸福地生活在牛奶瓶子里,靠腐烂的香蕉活着,抽屉里也爬满了蟑螂。但是摩尔根泰然自若地忙碌在这个肮脏的环境里。他坐在中间的一张桌子旁,他的主要武器是一个放大镜,详细观看是否出现了胡戈·德弗里斯所说的突变。如果某一个奶瓶里的果蝇没有出现他希望看到的突变或类型,他就用拇指把这些果蝇碾死,然后把它们的尸体随意抹在什么地方,例如在他的笔记本里。
1910年5月,戏剧性的突变出现了——一只果蝇的眼睛变成了白色,而原本所有的果蝇都是红眼睛。
摩尔根把白眼睛隔离开来,培育出很多白眼睛。他用白眼睛雄性果蝇与红眼睛雌性果蝇交配,结果很复杂。他用不同的方法培育各种后代,出现了一个激动人心的结果:他发现红眼睛与白眼睛的比例是3∶1。摩尔根听说过一个名词——基因,摩尔根还听说哺乳动物的染色体中雄性是X与Y,雌性是X与X,这里有三个X与一个Y。这可能说明,摩尔根发现了比例为3∶1的基因?
1900-1940年,达尔文理论陷入了黑暗时代。
这段时期,基因的情况也好不到哪里,全世界没有几个人说得清楚什么是基因。
染色体的情况更糟糕。染色体和核苷酸的拉丁名称又长又难记。行星是按照希腊神明的名字命名的;化学元素是按照神话、英雄和伟大城市的名字命名的……当时不受重视的23对46个染色体的名称,则干脆是按照它们的长度命名的:1号染色体的长度第一,2号染色体的长度第二……这些枯燥无味的命名,说明人们当时搞不清楚染色体和DNA的真实意义,所以对它们也没有什么兴趣。
非常幸运的是,只有一个重要的例外,就是果蝇。
摩尔根的团队不断推出新的发现,演示染色体的遗传效应,他给这些不同模样的果蝇分门别类,起了很多标新立异的品种名称。摩尔根想出了一个理论——复合基因(multiple genes)理论。他把果蝇的眼睛、翅膀的形状、绒毛的模样等作了分门别类,然后分别培育这些遗传特性。虽然基因和染色体没有改变,但是个体的遗传特性却变得互不相同,而且可以一代又一代培育出来。
虽然理论上没有什么新发现,但是,谁都无法否认摩尔根培育并展示出与性染色体有关的、不同类型的大批果蝇存在的事实。摩尔根变得越来越著名。
摩尔根的小小的纽约实验室原本拥挤狭小得滑稽可笑,1928年,成为生物学的“重要人物”之后,他搬到了加利福尼亚州的洛杉矶的宽敞明亮的新实验室里,雄心勃勃地希望建立自己的理论体系,虽然他的果蝇实验和突变理论实际上只是追随别人的实验模式和理论。他在洛杉矶加州理工学院(California Institute of Technology)创建的生物系,先后培育出了7个诺贝尔奖得主。
1933年,摩尔根获得诺贝尔奖。他是世界上第一个获奖的遗传学家。一个历史学家这样评述摩尔根:“他(摩尔根)建立了一些他曾经打算推翻的遗传学原理。”因为摩尔根的果蝇实验发现,遗传特性“往往不是受一个基因的影响,而是受多个基因的影响”。
事实上,当时诺贝尔奖委员会也不知道人类的染色体有多少?在给摩尔根颁发诺贝尔奖的时候,人们普遍认为人类有24对48个染色体。这个错误观念持续了大约30年。
1955年12月22日是一个具有历史意义的时刻,蒋有兴(Joe Hin Tjio,1919-2001,出生于荷属东印度(现印度尼西亚)爪哇的一个华裔家庭)最终发现和确认了人类有23对46个染色体。
1910年开始,赫尔曼·马勒(Hermann Muller,1890-1967)常常到纽约摩尔根的果蝇实验室参与一些工作。当时他是一个大学生,比摩尔根小24岁。后来马勒也成为一个著名的遗传学家,他用X射线诱导产生了基因的突变。
因为果蝇实验使人们知道,染色体和基因可能与生物遗传特性的变化相联系。马勒认为:孟德尔和达尔文的学说,互相完美地巩固了对方。马勒最终使得摩尔根也相信了这一点,最后摩尔根成为一个达尔文主义者。马勒强调,基因的作用大于自然选择。
虽然果蝇实验非常简单,但是在1920-1930年,这种办法成为一种国际流行的动物遗传学的实验模式。马勒回忆摩尔根的影响时说:“我们不会忘记摩尔根,他的例子影响了所有其他人,他的不屈不挠、深思熟虑、开朗和勇气。”
生物学家开始重新审视他们对达尔文的偏见,开始综合孟德尔的基因理论与达尔文的自然选择。生物学家发现,一些微小的变化确实可以改变物种演化的新方向。
果蝇实验和染色体基因作用演示说服了一些人,但是还有很多人不相信基因。在1940年DNA与蛋白的实验报告中,只有少数科学家认为DNA是遗传物质。
1952年,更好的证据出现了。阿弗雷德·赫希(Alfred Day Hershey,1908-1997)和他的女助理玛莎·蔡斯(Martha Cowles Chase,1927-2003),在美国纽约的冷泉港实验室(Cold Spring Harbor Laboratory)利用病毒进行的著名的赫希——蔡斯实验(Hershey-Chase experiment),证实了DNA是遗传物质。
蛋白中只含有硫,不含有磷。如果基因是蛋白,受感染的细胞里应该有硫。赫希和蔡斯在受到病毒感染的细胞里只发现了磷。也就是说,基因在DNA里,基因不在蛋白里。
赫希和蔡斯的这个实验激起了研究DNA的热潮。
1953年,一年之后,剑桥大学的两个年轻人,詹姆斯·沃森和弗朗西斯·克里克终于搞清了DNA的奇特而稳定的化学分子结构。1954年,20个青年学者(代表20种氨基酸)组成RNA领带俱乐部(RNA Tie Club),讨论分析DNA→RNA→蛋白的遗传关系:DNA是双链,RNA是单链,DNA将遗传信息交给“信使”RNA,然后由RNA指令细胞制造蛋白。这个遗传信息的转达和表达过程,转瞬即逝,机理难以查明。DNA→RNA→蛋白的遗传制造过程中,当然也会出现错误,但是细胞通常会立刻修正这些错误,否则这些错误就永久留在DNA里遗传下去。
(欧文·查戈夫(Erwin Chargaff,1905-2002)发现DNA的四个碱基比例为:A=30.9%,T=29.4%,G=19.9%,C=19.8%。这种比例明显意味着DNA碱基结构可能是对称的。1950年,他发表了自己的成果。1952年,查戈夫当面向沃森和克里克解释他的发现,帮助他们理解并最终搞清楚DNA的结构)
原子弹也无法改变DNA
1945年8月6日,广岛的幸存者们都记得原子弹的爆炸和火焰之间有一段奇怪的延迟,强烈的闪光过后是冲击波,静静地升起的蘑菇云越来越大越高,伽玛射线正在辐射着广岛的人群……遗传学家开始研究核辐射对DNA的作用。
1940年代,科学家已经发现基因存在于DNA的一些证据,但是人们仍然相信蛋白质才是遗传物质。科学家们继续深入研究,发现DNA和蛋白质之间存在特殊的关系。如果是DNA在指导制造蛋白,那么,核辐射是否会摧毁DNA?
1946年,马勒获得诺贝尔奖之后告诉《纽约时报》:“现在可以预言1000年以后的结果,那些被原子弹杀死的人比他们(原子弹幸存者)更幸运。”
马勒是一个悲观主义者。
但是,遗传学家在幸存者中根本找不到伽玛射线损害DNA的证据,几年甚至几十年以后也没有发现证据。如此强烈的伽玛射线,对DNA竟然没有伤害?虽然幸存者们经历着各种折磨,很多人在几十年后陆续死于核辐射后遗症,但是,DNA确实没有改变。
细胞修复DNA双链的速度,超过简单复制DNA时的速度大约3 000倍。长度大约2米的DNA缠绕压缩在染色体里,当原子弹的伽玛射线打击这些双链时,如果其中一个链条受到损害,细胞可以立刻从另一个链条复原两条双链:因为DNA的两个链条是互补对应的。如果无法复原DNA,细胞就会通过自杀来实现自我牺牲。如果同时自杀牺牲的细胞太多,人就会死亡。
虽然伽玛射线可以造成幸存者们出现皮肤溃烂、脱发、体虚、咳血等症状,但是这些都会逐渐恢复。只是那些没有被杀死的细胞发生基因突变的可能性增大,而这些突变的累积可能导致癌症。日本随后出现的很多白血病(血癌)病例证明了这一点。10年之后,日本的白血病高潮才逐步消退。
原子弹受害者的孩子们也没有受到影响,这进一步证明了DNA没有改变。
很多科学家的悲观预测都没有出现,氢弹之父爱德华·泰勒(Edward Teller,1908-2003)甚至猜测少量的原子辐射对人可能有正面影响。总而言之,原子弹的力量似乎无法影响DNA的遗传。
(西德尼·布伦纳(Sydney Brenner,1927-),罗伯特·霍维茨(Robert Horvitz,1947-),约翰·苏尔斯顿爵士(Sir JohnE. Sulston,1942-),2002年,这三位科学家获得诺贝尔奖。他们在细胞的凋亡(apoptosis)方面分别做出了贡献。细胞的凋亡是一种程序性死亡,是细胞主动实施的死亡)
山口彊(Tsutoma Yamaguchi,1916-2010)是一个典型的例子。1945年,山口彊在广岛和长崎两次受到原子弹辐射。在受到辐射的当时,山口彊的皮肤溃烂、头发脱落、左耳丧失听力。后来,他的所有症状都消失了,头发也长了出来。他又活了65年,2010年因胃癌去世,享年93岁。科学家们推测,山口彊的细胞修复DNA和RNA的能力特别强。
DNA的强大,令科学家们感到不可思议。暴露在宇宙射线和太阳辐射之下30多亿年的DNA,虽经历过无数的损害,却始终没有改变。大自然怎样赋予DNA不可思议的保护模式?我们至今对此一无所知。科学家仅仅知道遗传过程是DNA→RNA→蛋白的简单遗传教条,迄今为止,任何人都不知道真正的DNA的“语言”和“数学”模式。
地球上的生命形式诞生于约35亿年前,所有有机组织都采用了同一套DNA体系进行生命的遗传。最近的4亿多年里,地球曾经发生过五次生物大灭绝,大体情况如下:
第一次约4.4亿年前,奥陶纪末期:约85%的物种灭绝。
第二次约3.6亿年前,泥盆纪后期:海洋生物遭受灭顶之灾。
第三次约2.5亿年前,二叠纪末期:海洋95%和陆地75%以上的物种灭绝。
第四次约2亿年前,三叠纪晚期:爬行动物遭遇重创。
第五次6 500万年前后,白垩纪晚期:侏罗纪以来统治地球的恐龙灭绝。
DNA的强大和不可改变,至今无法找到解释。
每一个人的身上,都带着很多遗传错误,但是大都是非致命的错误。所有的生物组织,随着年龄的增加都会继续积累突变(错误):每个人的细胞平均大约每天复制50万次以上,有的细胞每天复制150万次以上,在人的大约100万亿个细胞的复制过程中,想避免错误非常困难。生物组织形体越小、外界温度越高、细胞的DNA复制越活跃,发生复制错误的概率越高。人类属于体形较大的哺乳动物,体温也基本保持着恒温,所以人类的细胞复制基本保持恒定的速率。
两种DNA的发现与解释
沃森和克里克破解DNA的双螺旋结构,与一个女科学家密切相关。
罗莎琳·爱尔西·富兰克林(Rosalind Elsie Franklin,1920-1958)是英国生物物理学家、X射线专家,对发现DNA、RNA、病毒、煤炭和石墨等的分子结构做出了重大贡献。她在英国伦敦国王学院(King’s College London)工作期间,与同事莫里斯·威尔金斯(Maurice Wilkins,1916-2004)合作,第一个做出了DNA的X射线衍射图像,初步揭示出DNA的结构。
1962年,她的同事威尔金斯以及沃森和克里克等三人分享了1962年的诺贝尔奖,他们发现了DNA的双螺旋结构和遗传的机理。当时,罗莎琳已经去世4年了。很多人评论说,罗莎琳也是应该获得这次诺贝尔奖的科学家。
沃森和克里克搞清楚DNA的基本结构之后,另一位女科学家对DNA的子结构——碱基的结构做出了贡献。这个人就是米利亚姆修女(Sister Miriam Michael Stimson)。
米利亚姆修女1939年开始研究DNA的结构,1953年她获悉沃森和克里克的研究结果后,知道自己十几年的心血白费了。但是米利亚姆修女没有放弃,转向了研究DNA的碱基的结构和其他领域。
1953年,沃森和克里克发表DNA结构的同一年,米利亚姆修女在索邦大学(La Sorbonne,巴黎大学体系的一部分)发表了她的研究成果。这是继居里夫人之后,第二位妇女走上这个法国最高讲坛。2002年,米利亚姆修女以将近89岁高龄去世。
科学的历史上充满了重复发现现象:自然选择、氧气、海王星、太阳黑子……两个、三个甚至四个重复的独立科学发现同时出现。
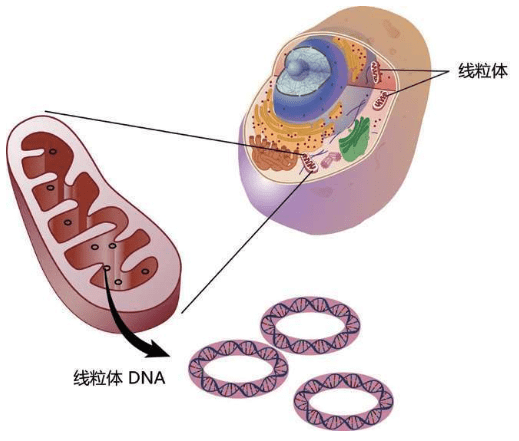
1963年,两个研究团队几乎同时发现了人类存在第二个DNA的重要事实。一个团队在显微镜里看到线粒体,形状像豆子的组织,在细胞里提供能量;另一个团队把肠子煮成浓汤之后提炼出线粒体。两个团队都发现,线粒体有自己的DNA。19世纪,DNA的发现者米歇尔并不知道有两个DNA,他以为DNA的唯一住所是细胞核。
现在看起来,当时科学家们对线粒体DNA的解释非常幼稚可笑:这些DNA是从细胞核“借用”的DNA,但是用完之后没有归还给细胞核。
生物科学的历史,总是被不断改写。3年以后,1966年,一位美国女科学家对线粒体DNA作出了颠覆性的解释。
1966年,琳·马古利斯(Lynn Margulis,1938-2011)在她的论文中彻底颠覆了人们对线粒体DNA的好奇心,把人类的视野推向了一个全新的高度。马古利斯认为:动物——植物——菌类都起源于原生生物,生命不是通过互相战斗而是通过互相协作占据了整个地球。
马古利斯认为,地球上所有的生命都分享某些基因,我们所有生命都是地球上第一个微生物的后裔。很久很久以前,一个大的微生物吞下了一个小的微生物,不知道经过多少世代之后,双方形成共存模式:大微生物不再生产能量,只是提供庇护和原料营养,小微生物则负责用氧气生产高辛烷值的“燃料”能量。
正如哲学家亚当·斯密(Adam Smith,1723-1790)当年的预测:这种生物分工形成互惠互利的关系,双方失去对方都会导致死亡。
这就是线粒体的起源。
马古利斯的理论已经完全超越历史上所有的传统生物学家的梦想,也得到越来越多的来自各个学科的证据支持。线粒体DNA的存在,只是内共生学说的证据之一。
但是,人们对这个理论的接受,却并非一帆风顺。马古利斯的这篇文章曾经被大约15家杂志退稿,最终发布以后,很多科学家拒不接受这个理论。他们每一次攻击马古利斯,马古利斯都拿出新的一系列证据予以反驳。双方阵营的辩论规模越来越大,对手们批评马古利斯的理论虽然完美,唯独缺乏证据。越来越好斗的马古利斯干脆对观战的无数听众直接发出呼吁:“这里有生物学家吗?有没有分子生物学家?”她伸开双手大笑着问道,“是的,我知道你们讨厌这个(内共生学说)!”
马古利斯说对了,生物学家们确实讨厌内共生学说。
这场争吵持续了十几年,直到1980年代发明新的扫描技术之后,人们终于发现,线粒体里存放的DNA不是长长的线性的染色体(动物和植物都是线性的长形的DNA),而是环形的DNA——只有细菌才是环形的DNA染色体。线粒体确实是一个古代细菌。
线粒体里有37个类似细菌的蛋白,它们的A-C-G-T序列与细菌非常相似。马古利斯预测的证据找到了,科学家们甚至找到了线粒体DNA的活着的亲戚,例如伤寒细菌(typhoid bacteria)。(在34.65亿年前的岩石中发现过一个类似微生物的多单元细丝(A microbe-like cellular filament foundin 3.465 billion year old rock),这是地球生命起源于35亿年前的证据之一)
各种证据接踵而来,反对者们被批驳得哑口无言。马古利斯的理论不仅解释了线粒体,还帮助理解了地球生命的深奥秘密——为什么在生命发生之后,演化的速度达到失速的状态。没有线粒体的推动,原始的生命不可能演化为高等生命,直到出现人类。
我们不知道原始的生命元素来自哪里?也许在海洋底部的火山口,也许在外太空。天文学家已经发现外太空星际尘埃中漂浮着多种氨基酸,化学家也测算出DNA的碱基可以在空间形成。外界环境的恶劣,反而导致更加复杂的分子的形成。有的科学家认为,也许是彗星带着这些原始生命元素落入了海洋。它们组合和演化的过程非常缓慢,不知道经历了多少年。也许很多年里根本没有什么演化,只是处于一种蓄势待发的“孵化”状态。
所有生命的共同起源出现之后,很短时间就分化了。不同的生命,采取了不同的消费能量的模式。原始的微生物,可能仅仅需要2%的能量复制和维持DNA,但是要花费75%的能量用于蛋白质制造DNA的过程。所以,如果一个微生物能够找到另一个寄生的微生物负责生产能量,则是一个意义巨大的进步。这样,简单的生物很快就会演变成极其复杂的多功能的新生物——线粒体,可以使细胞生产DNA的能力激增20万倍,这在过去的演化史上几乎难以置信。
马古利斯的理论,撕开了演化史中黑暗的内幕:人类根本不需要那么多营养。
线粒体DNA的真相,为科学打开了一个新的世界。人类正是利用它最后追寻到了17万——20万年前的“夏娃”。
马古利斯的威望日盛,她的理论使得达尔文和孟德尔的理论成为“配角”,有人称其为新达尔文主义。马古利斯认为,各种生物都是合成的,例如美人鱼、斯芬克斯等,虽然目前还没有证据。马古利斯得到了两个对立阵营——激进的和保守的生物学家们共同的赞美,因为这种理论包罗万象、极其完美,引起了各种新的研究和探索,各种观念都已经或正在发生变化,例如人们现在认为生命不应该用名词Life,而应该用动名词Living。
更加重要的是,这个理论改变了人们对人类——地球——生命的认识,改变了对宇宙的认识。我们必须控制发展、保护环境,走可持续发展、绿色能源的道路……
人类还发现,原来的生物分类的思考方向也错了,至少并不全面。地球的生命类型如果从利用二氧化碳和氧气来区分,可以分成两大类:
换句话说,人类原本就是“寄生类”生物,更不应该打破这种生态平衡。最近一万年来,过度贪婪的农业文化不仅正在打破这种生态平衡,甚至改变了大气温度和地球环境。如果继续向地球无限制地索取,向大自然的整体生态系统挑战,人类最终的失败将是毫无疑问的。
地球上的所有生命,都有一个共同的微生物的先祖,如下图所示:
这是一棵生命树,所有的生命都是地球上第一个微生物的后裔。人类属于异养生物,必须依赖食用其他植物、动物和菌类等才能生存。
国际专家曾经多次组织研究小组,确定了若干个对人类生存至关重要的“地球生命支持系统”,并对目前人类的消耗水平和系统的“临界点”进行了量化和评估。科学家们警告:一旦这些临界点全部或大部分被突破,人类生存环境将面临不可逆转的变化。
这些地球生命支持系统包括但不限于:
海洋酸化 臭氧浓度 淡水消耗量
生物多样性 氮磷循环 土地使用率
二氧化碳浓度 气溶胶浓度 化学污染
大自然不遵守大自然法则
1909年,一个农民带着一只鸡来到位于纽约长岛曼哈顿的洛克菲勒大学(Rockefeller University)。这所大学当时名称为Rockefeller Institute,是石油巨头洛克菲勒捐赠创立的私立大学。这所大学先后诞生了24位诺贝尔奖获得者。
这只鸡病了,它的右侧胸部长出可疑的肿瘤。当时这种肿瘤正在肆虐美国,很多农民因为可能损失养鸡场而焦虑不安。
佩顿·劳斯(Peyton Rous)负责诊断这只鸡的肿瘤。他没有治疗这只鸡,而是杀死这只鸡,提取出含有肿瘤细胞的液体。劳斯估计,肿瘤细胞里可能含有某种微生物。他把这种液体注射到另外一只鸡的胸部,第二只鸡很快也出现了肿瘤。
劳斯继续重复实验,六个月里,一个又一个鸡肿瘤出现了。
劳斯困惑了。他在霍普金斯医学院学习时,知道病毒可以传染疾病,但是肿瘤不会传染。但是他的实验只有一个结论——病毒在传染肿瘤。他无法相信这个荒谬可笑的结果,但是,他还是发表了这个结论。
劳斯的发现,用现在的术语表达就是病毒通过RNA传染癌症。
科学界很快忘记了劳斯的这个论文。虽然后来也有人偶尔发现病毒与肿瘤之间的联系,但是其他新发现很快又掩盖了这类发现。但是人们对于病毒的原理也越来越清楚——它们利用细胞制造它们自己。
1958年,DNA结构的两个发现者之一,弗朗西斯·克里克(Francis Crick)发布了著名的“分子生物学中心法则”(Central dogma of molecular biology)。这个中心法则的主要含义是:DNA制造RNA制造蛋白质(DNA makes RNA makes protein)。
显然,劳斯错了,RNA病毒重写细胞违反分子生物学中心法则。
非常遗憾,1960-1970年的一系列发现证明:大自然不遵守大自然的法则——大自然根本不在乎什么分子生物学中心法则。支持克里克的其他诺贝尔奖获得者也都错了。生命的形式绝不是那么简单,那么绝对,那么遵循“大自然法则”。人类对于生命,仍然一无所知。
例如,病毒的逆反方向翻译和复制也是可能的,HIV病毒就是一个最好的例子。深谙遗传技术的HIV病毒可以熟练地操控DNA,哄骗被感染的细胞把病毒DNA塞进细胞的基因组。也就是说,这些HIV病毒把细胞搞糊涂了,不知道制造的是“他们的”DNA还是“我们的”DNA。于是,HIV这类RNA病毒摇身一变成为人类的DNA,患者的艾滋病(AIDS)于是变得不可收拾——人类的免疫系统搞不清楚马奇诺防线的哪一边是敌军了。
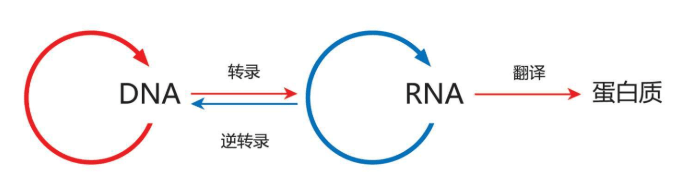
很多科学家猜测,在演化历史上,RNA出现的时代比DNA更早。
1986年,另一位诺贝尔奖获得者沃特·吉尔伯特(Walter Gilbert)首先提出“RNA世界假说”(RNA world hypothesis),因为RNA可以自己制造自己。这个假说得到很多科学家的支持,其中包括另外几位诺贝尔奖获得者。
这些理论得到了继续演化,地球从RNA世界逐步演化到今天的“DNA-RNA-蛋白质世界”(DNA, RNA and protein world)。在这个新的世界里,结构稳定的双链DNA最终“降伏”了活泼好动的单链RNA,作为自己的信使传播制造蛋白的遗传信息等等。
荷马史诗不能总是口头流传,必须经由“稳定”的文字记述才能世世代代流传下去。DNA就是荷马史诗的永恒不变的文字内容,RNA多才多艺、声音惟妙惟肖、姿势活龙活现,大家一起保留了历史。
如果顺序搞反了,细胞制造的就不是原来的DNA,而是病毒的DNA。1909-1911年,劳斯发现的正是这种病毒之一。这些病毒被注射进鸡的体内以后,它们“欺骗”鸡的细胞进入核DNA,然后复制的都是这些病毒的DNA。最后复制速率不可控制,疯狂地增殖形成恶性肿瘤。这种通过病毒传染的肿瘤比较少见,大部分恶性肿瘤的产生源自基因突变的累积。
劳斯的论文在当时默默无闻,整整55年之后,他的发现才得到各种新证据的证明。这种感染癌症的病毒最后以他的名字命名为劳斯肉瘤病毒(Rous sarcoma virus)。这种病毒正是逆转录感染:先从RNA感染DNA后,再返回来制造蛋白,然后不断繁衍复制。
当时,劳斯也不明白为什么病毒会传染癌症?他猜测也许病毒释放某种毒素导致癌症。但他坚信自己的研究,他的坚持在55年后得到公认。
1970年,获奖4年后,劳斯去世,享年90岁。他去世以后,被大批年轻的微生物学家视为崇拜的偶像。劳斯因为一只鸡获得诺贝尔奖,他也是等待诺贝尔奖时间最长的一位科学家——整整55年。
2000年,人类基因组工程完成以后,生物学家发现:不到2%的基因是编码遗传基因(即编码制造人类的蛋白质);超过8%以上的基因是病毒细菌的基因。
超过98%以上的非编码基因作用不明,有待继续研究;换句话说,细菌病毒基因数量是人类遗传编码基因的整整4倍以上。所以很多生物学家开玩笑说:“人类基因组工程是不是命名不当?”
这是怎么回事?
从病毒的角度来看,殖民到动物的DNA是理所当然的。很多病毒狡猾奸诈不择手段,通过逆转录导致癌症或疾病,例如艾滋病毒(AIDS)。但是,并非所有病毒都是害虫。大多数病毒都心怀慈善、自我约束,绝对不做伤害自己宿主的愚蠢事情。有一些深谋远虑的病毒干脆进入宿主的精子或卵子,随之进入宿主的后代的体内,世世代代存在下去。还有一些无害的病毒,甘当宿主的对敌作战“工具”——宿主可以直接利用它们打击入侵者,它们也便成为人类免疫系统的成员。这个道理其实非常简单:赌场喜欢雇用精明的发牌员,电脑保安公司喜欢雇用黑客高手,因为他们知道如何战胜对手。我们进入农业时代之后,又获得很多其他病毒作为开关,比如放出一些酶,消化各种小麦、玉米等碳水化合物。
在我们体内,这些病毒、细菌、原生生物基因无处不在,操控着人类演化的方向盘,它们构成的无数开关和调节器控制着人体的活动和大脑的思维。
基因的变化并非只有这些:人类DNA的大约一半是机动DNA(mobile DNA)或曰跳跃基因(jumping DNA),有的转位子基因(transposon,又称Transposable element)的长度甚至达到300个碱基。这些基因的位置和功能是可以转换的。首先发现这些基因特性的芭芭拉·麦克林托克(Barbara McClintock,1902-1992)最初不被人们接受,直到32年之后,她才得到承认。1983年,麦克林托克获得了诺贝尔奖。
那么病毒细菌基因是否仅仅占人类基因组的8%呢?现在还不清楚。很多科学家承认:“我们对于我们人类这一物种的整体认识,可能都错了。”
地球是一个活的超级生物体
在古希腊神话中,最高的神明是盖亚(Gaia)。我们必须了解盖亚理论,才能理解生命的起源和人类六万年的旅程。
在希腊语中,盖亚(Gaia)是地球女神或地球的拟人化,她是所有生命的起源,她是所有提坦(Titan)和巨人的母亲,也就是说是地球孕育了生命。因此,盖亚也被西方人用来代称地球。
首先,我们看一看两个问题及其答案:
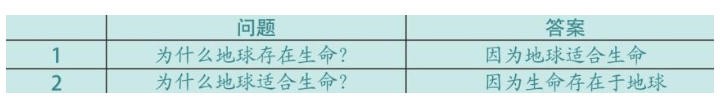
上面两个问题及其答案,互为依存,互为因果。这是古希腊柏拉图时代提出的问题,这也是现代科学理论——盖亚理论(Gaia theory)的由来。即地球上的生命本身产生了巨大的反馈,确保了自身的生存条件。
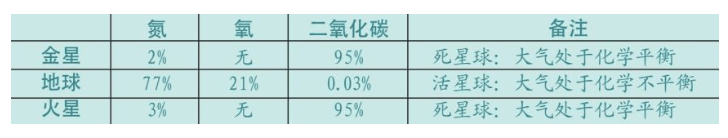
现在,让我们看一看地球旁边的两个星球——金星和火星的条件。金星比地球靠近太阳,火星比地球远离太阳,但是地球两边的这两个星球都不适合生命存在。
地球上的这些“生命存在的条件”都是生命体系在几十亿年里自己创造出来的。
那么,生命是什么?
生命是地球生命的集合,是所有地球上生物的集体行为,从最小的细菌到最大的哺乳动物,都在协同维持地球的最佳环境。换句话说,地球生命的活动,正是为了确保自己的生存。
盖亚理论设定地球为一个完整的整体,一个超级生物体,维系着地球上所有生命形式的存在。盖亚理论强调了一个事实:我们所有生命都是互相关联的,每一个物种的生存都与我们自己本身的生存息息相关,任何一个物种的灭绝都会给地球带来灾难。
20世纪后半叶,我们见证了对于地球的理解的不可思议的大飞跃。
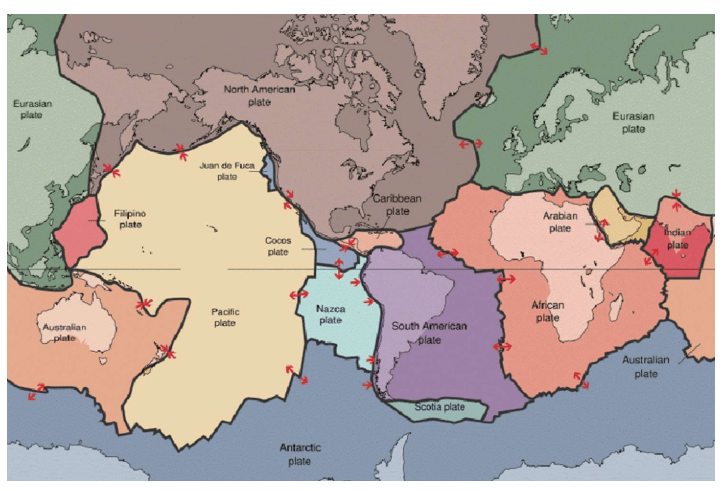
1910年代,魏格纳(Alfred Lothar Wegener,1880-1930)提出了不可思议的地球大陆板块漂移学说。这个理论经过很多学科的共同研究,最终得到确认。(大陆板块漂移说另一个发现者,是赫斯(Harry Hammond Hess,1906-1969))
1960年代,詹姆斯·洛夫洛克(James Lovelock,1919-)最早归纳整理出盖亚假说。这个假说认为地球是活的。
1979年,洛夫洛克正式发表盖亚假说。假说认为地球的物理——化学——地质——生物过程都是互动的,生命和非生命组成了地球。“盖亚”是温和的、多产的,也是无情的。洛夫洛克提出地球处于一种动态平衡状态,维持相对稳定的条件,地球本身就是一个生命体——大气层、海洋、陆地、生物圈等构成太阳系里的一个有机组织。洛夫洛克把这种观点叫作地球生理学(Geophysiology)。这种观点,其他星球也同样适用。
1965年,洛夫洛克在美国航天局(NASA)参与寻找火星的生命时,产生了“存在盖亚”的思想。当时,他被要求提出一些“其他星球可以存在生命的假设”,他提出的其中一个假设是大气层的化学平衡:如果一个“死”星球的大气层是化学平衡的,也就是说,如果所有可能发生的化学反应已经发生过了,那么,这个星球的大气层的气体是相对惰性的。换句话说,如果生命存在于某一星球上,这个星球的大气层就不是平衡的,化学反应会非常活跃地持续进行。根据这个假设,洛夫洛克和他的美国航天局团队研究了火星和金星上的大气成分,发现这两个星球上的大气层主要由通常不再进行化学反应的二氧化碳构成,它们分别占火星和金星大气层的95%,所以这两个星球是“死”的。但是,地球的大气层与众不同,属于不稳定的多种气体的混合,所以,在地球上可能存在生命。而事实上,我们地球确实存在生命。
“地球是活的”这个理论曾经使很多科学家感到非常苦恼,但是,仅仅20年后,盖亚假说就得到承认。虽然盖亚假说还要接受时间的检验,但是已经颠覆了我们对地球的认识。
洛夫洛克在他的第一本书的第一章结束时写道:
如果盖亚是存在的,她和人类的关系就是非常重要的,因为人类是这个复杂的生命系统中占据支配地位的动物物种,有可能左右各种力量之间的平衡……
有的人喜欢在地球上走路,有的人喜欢站在地球上,有的人喜欢凝视着地球,他们都为地球上承载的如此绚丽多彩的生命而惊叹不已……盖亚假说为“人类为什么出现在地球上”给出了一种思考模式——与大自然是一种冷酷无情的不可战胜的力量的悲观观念相比,盖亚假说给出了另一种选择……
但是,盖亚假说也给出了同样令人沮丧的另外一幅画面,因为这个星球似乎像是神经错乱的一艘宇宙飞船,围绕着太阳的内核在永不停息地飞行着,既无人驾驶,也没有目标……
这就是盖亚假说的来由,这个过程也告诉我们科学家是如何思考和研究的。
地球大气成分不是化学平衡的,但是仍然维持着一种稳定状态,说明地球大气层存在某种形式的行星调节机制。洛夫洛克猜测,可能生命本身维持着大气层的成分,后来他把这一概念拓展到涵盖气候——岩石——空气——海洋的整个体系,这个体系形成了一种自我调节过程。
(1979年洛夫洛克出版了《盖亚:对地球生命的新看法》(Gaia: A New Look at Life on Earth),这是洛夫洛克出版的“盖亚理论”的第一本书。他出版了多部著作,如:
Lovelock, James.
Gaia: A New Look at Life on Earth.
Oxford University Press, Oxford, England.
洛夫洛克的这个观念,其实并非全新的观念。)
世界著名的“地质学之父”詹姆斯·赫顿(James Hutton,1726-1797)最早提出“地球是活的,地球是一个超级生物体”这一观念。
1974年,在洛夫洛克发表他的盖亚理论之前,另一位医学博士兼著名作家刘易斯·托马斯(Lewis Thomas ,1913-1993)在他的著名随笔《一个细胞中的生命:一个生物学观察者的笔记》(The Lives of a Cell:Notes of a Biology Watcher)里写道:
如果在遥远的月亮上观察地球,就会吃惊地发现地球是活的。月亮的表面是干巴巴的,四处坑坑洼洼,就像一块干枯的骨头。在月亮的上方,正在自由自在地升起一个潮湿的、亮闪闪的地球,蓝色的天空就像是地球的细胞膜,这是宇宙中仅有的一个星球。如果再仔细观察,就会看到巨大的白云形成的涡旋,覆盖了大约一半的陆地。如果你以一个地质学的时间跨度观察,就会看到各个大陆本身也在移动,好像一些内在充满热情的外壳板块。地球是一个有机物,本身就像一个活着的生物,满载着信息,被太阳熟练地操控着。
刘易斯·托马斯进一步写道:
我曾经试图假想地球是一种有机组织,但是它不是,地球更像是一个细胞。
这个令人震惊的理论让我们知道,所有生命形式都是内在关联的,都对与每一个人、每一个生命息息相关的环境的维持作出了贡献。是所有生命的行为的总体协同构成,帮助地球维系着一种令人惊异不已的“活的星球”的环境质量。
我们人类曾经认为自己是地球上的特殊物种,真正的智慧物种,神造的物种。但是,盖亚理论打开了我们的视野,揭示了地球的真相:我们人类和所有其他生命,都是地球这个超级生物组织的组成部分之一。
这一事实使我们认识到,无论我们的科学和技术如何绚丽夺目,我们仍然被包裹在这个超级生物组织——地球之内,不论我们是否情愿,我们仅仅是比我们更加伟大的世界的一个部分。
如果说,地球上存在着真实的生命,这个生命就是地球本身。其他所有生命形式都是转瞬即逝的。假设我们坐在外太空的一把椅子上,观看地球围绕太阳运转,能够看到生命吗?假设我们以六万年为尺度观看地球生命,1分钟等于1 000年,能够看到生命吗?我们看不到连续存在的任何一个生物,包括人类,所有人都转瞬即逝,迅速消失于无形。只有唯一的一个生命体持续存在,那就是地球。
只有地球,六万年里始终支持着所有生命形式的活动,化学的和物理的形式的活动。天空、湖泊、海洋、山脉、花朵、小鸟、大型哺乳动物、你、你的朋友、整个人类都是活生生的地球的组成部分而已。我们所有人的贡献,只是维持这个地球生命的可持续发展。这个世界上的一切都是互相依存的。根本不存在什么至高无上的概念。在“地球盖亚”面前,所有的生命都是平等的,都是地球的一部分。
不论“有机组织”“细胞”还是“超级生物体”,都仅仅是语义表达的差异。从哲学角度,盖亚理论的关键在于:地球是一个单一系统,设计合理,自我调节,物理、化学、地质、生物的力量综合互动,作为一个完整的整体维持着来自太阳的能量输入,和散发到外太空的能量输出之间的平衡。
地球的基本配置正是调节能量流量和物质循环:

盖亚假说的“地球作为一个单一的系统而行为”的理念带给我们全新的警示:
这个星球上的所有事情都是相互关联的,人类的行为处于一种全球过程中。我们不能继续认为地球上距离遥远的事物与己无关,我们不能继续认为这个星球上的某一行为是独立无关的。这个星球上发生的每一件事情,无论砍伐树木还是植树造林,无论增加还是减少二氧化碳排放,无论开垦耕地还是退耕还林,都会影响整个星球。
甚至微观生命系统的设计也是极其精妙的。2008年,美国《科学》(Science)杂志发表了一篇在肯尼亚实验保护金合欢树的研究论文。实验将一半金合欢树用栅栏围住,阻拦大象和长颈鹿等食草动物,另一半完全放开,放开的金合欢树将承受着极大的来自食草动物的压力。实验结果与人们的预测完全相反,被大象和长颈鹿蹂躏的金合欢树郁郁葱葱,而被保护的金合欢树却越来越虚弱、濒临死亡。
研究发现,金合欢树有一种防卫体系,其中包括一种特殊小蚂蚁。金合欢树分泌含糖汁吸引和滋养这些小蚂蚁,被栅栏保护以后,树不需要自我保护,就停止分泌含糖汁,这种特殊小蚂蚁也不来了,换成另外一些蚂蚁品种,其他昆虫和害虫乘机攻击这些金合欢树,于是它们日益衰败。这个简单的例子说明,在一个相对容易了解的生态体系里,每一种生物组织的交互关系都是错综复杂、难以预测的,当我们试图对某一元素修修补补的时候,就存在着其他关联元素的后续效应风险。
如何理解“地球是活的”这一概念呢?首先,让我们看看科学家如何理解生命。
物理学家定义生命是一个局部减少熵的系统(生命与熵的战斗)。从分子生物学家的角度来看,生命是复制DNA达成生存并根据周围环境的改变而演化以达成最佳生存。生理学家认为,生命是一个生物化学系统,利用外来的能量而生长并重复生产。
对于地球生理学家(Geophysiologist)洛夫洛克来说,生命是一个向物质和能量开放的系统,同时维系着一种内在的稳定状态。
加利福尼亚州的红杉(S e q u o i a gigantea)是生命的最好注解。这些巨树生长在树丛里,高度达到100米以上,寿命超过3 000年。红杉97%的组织是死的,主干和树皮已经死去,只有主干外表的细胞部分是活的。红杉的主干类似地球的岩石圈,只有岩石圈外表薄薄一层生物圈是活的。红杉的树皮类似大气层,保护着这层生物圈,并且进行生物学意义上非常重要的气体交换——二氧化碳和氧气的交换。
毫无疑问,红杉总体上是活的生命,我们不能只把红杉的外层称为红杉,其余部分视为死的木头。
地球的很大一部分可以视为“非活的”(岩石圈),但是这些“非活的”地球部分也是生命过程的某种延伸,所以地球是活的,就像红杉。为了更好地理解地球的“生理”,我们对比一下人的体温和地球的温度调节机制。
人体温度维持在摄氏37度,这是大脑、人体组织、外界系统之间反馈的结果。我们的身体发展出提高和降低体核温度的一套感应器系统:如果太冷了,身体就会产生热量;如果太热了,身体就会散发热量。但是,人体的调节能力有限,所以发明了衣服隔绝过热或过冷的温度,衣服使人类可以生存在非常寒冷的北极或者最热的沙漠。
地球温度调节机制与此类似,但是地球的“衣服”复杂得多。首先,地球对太阳的光线有一种反照率(Albedo),反照率系指星球的颜色、吸收或反射光线的能力,比如沥青的马路和白色的路边小道的反照率就不一样。
地球调节温度的方式如下:
黑色区域:例如夏季的山脉、森林、海洋,可以从太阳那里吸收更多的热能。
浅色区域:例如沙漠、云区、极地冰盖,可以反射更多的热能。
地球的反照率不是恒定的,其中云团是调节全球温度最重要的因素之一。如果云团较多,从地球反射的阳光也多,地球温度就会下降;如果云团较少,更多的阳光抵达地球表面,地球温度就会上升。
那么,哪些因素在控制云团的多少?
控制云团的因素很多,大气层与海洋的互动是最重要的因素。
海洋占有地球表面的三分之二,海洋上形成的云团是地球温度最重要的一个因素。
最近几十年里,人类刚刚搞清楚海上浮游植物,尤其是coccolithophorids的云团冷凝核(cloud-condensation nuclei,CCN)机理。云带来雨是众所周知的常识,但是谁也不知道云在海上是如何产生的。
人们通常认为,阳光的热效应使海水蒸发成为云。其实,还有更重要的原因。大批小小的绿色浮游生物始终漂浮在海面上,它们的生命循环的一部分就是向大气中排放硫。正是这些硫分子“鼓励”水分子凝聚在一起形成云升上去,再形成雨滴降落下来。水蒸发到大气层之后进行浓缩和冷凝,但是,构成云团还必需一些小颗粒作为“核”来聚集水分而形成小水滴。这些小小的颗粒叫作云团冷凝核,存在于大气层里,它们形成了云团。这种物质叫作二甲基硫(dimethyl sulphide,DMS),也来自海洋中的浮游生物。近年的研究已经定量地确认了这些海上浮游生物如何排放二甲基硫进入大气层,然后如何形成了云团。也就是说,这些海上的小小绿色浮游生物群体不仅帮助形成云,还帮助形成雨,它们对整个地球的温度调节做出了巨大的贡献。
这些小小的单细胞生物是地球的恒温器:太阳光线比较强烈——浮游生物加速繁殖——产生大量二甲基硫——产生更多云团——云团遮挡阳光——降低地球温度——浮游生物繁殖减速——云团减少——阳光增加——地球温度升高——浮游生物再次加速繁殖……如此循环往复,地球的温度得以保持稳定。
这是盖亚理论中生物圈影响大自然(调控地球温度)的一个典型例证。
地球的岩石圈、大气层、水圈、生物圈构成了一个和谐的、有智慧的、有哲理的、富有诗意的整体。地球是一个活的生物组织,生物、地质、化学、水文学的过程都是关联的。
这个机制,诞生了“生物地球化学循环”(biogeochemical cycles)这个新名词。也就是说,地球的物质和化学元素必须出现物理的循环,如果这些物质和化学元素固定不动了,这个体系就停滞不变了,地球就会变得像月球一样死气沉沉。
最常见的生物地球化学循环是碳循环——氮循环——硫循环。这些元素的不断循环是活的生物组织的关键所在。大量的物质被活的生物组织消耗——转换——运送——再循环……这种生物组织控制的行星过程,使我们相信盖亚假说。大量证据出现以后,盖亚假说被称为盖亚理论。盖亚理论带来了哲学性的科学性,引发很多新的思考甚至新的学科,例如行星生物学(Planetary Biology)。
事实已经证明,地球的物理、地质、化学、生物过程确实是互相依存的。这种动态平衡与人体内的动态平衡概念类似。海洋和河流是地球的血脉、大气层是地球的肺、陆地是地球的骨骼、生物组织是地球的感官……对盖亚理论的研究似乎像研究一种“地球生理学”。
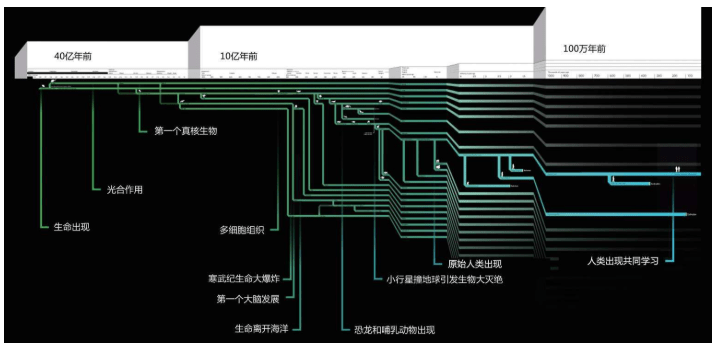
04:00:生命出现,厌氧生物。此后进展缓慢
20:30:出现第一批海洋生物,不久出现水母
21:04:出现三叶虫等复杂的海洋生物
22:00:陆地出现植物,接着出现动物
22:24:陆地被森林覆盖,成为今天的煤炭
23:00:恐龙出现,统治地球约40分钟后灭绝,哺乳动物的新生代开始
23:58:43:人类出现,至今已有1分钟17秒的漫长的进化历史
洛夫洛克和马古利斯认为:因为地球是活的,所以生命不是名词Life,生命应该是动词或者动名词Living。这个用词的变化,意义非常深刻。
反对这种“活的有机组织”观念的最主要论据是地球不可再生,而活的有机组织都是可以复制并把自己的遗传信息传承给后代。但是,生命正是盖亚诞生的,人类也正是盖亚诞生的,而所有这些生物都是可以再生的。
盖亚理论,无所不包,无法反驳。
盖亚理论最有意思的延伸之一是这一种理论转换成为多种理论。这种现象在科学领域的历史上极其罕见,说明盖亚理论蕴含的勃勃生机。盖亚理论正在向各个科技领域不断延伸拓展,每一个科学家及其信仰都受到巨大冲击。
盖亚理论现在已经得到公认,虽然仍然有少数人质疑这套理论,但是所有的反对者也都一致承认:生命确确实实影响着这个行星的各种体系的过程。
盖亚理论公认的两位创始人中,另一位创始人是美国女生物学家琳·马古利斯(Lynn Margulis),她第一次提出“生命或生物圈调节和维持气候和大气层的成分以维持其最佳状态”。这个解释,与洛夫洛克不谋而合。
马古利斯出版了很多论文和著述论述生命的起源,在世界科学界引发了一场又一场大辩论。最后,马古利斯赢得了胜利,世界的生物分类也作出了如下的改变:动物和植物两界,扩展为三域五界,外加病毒。
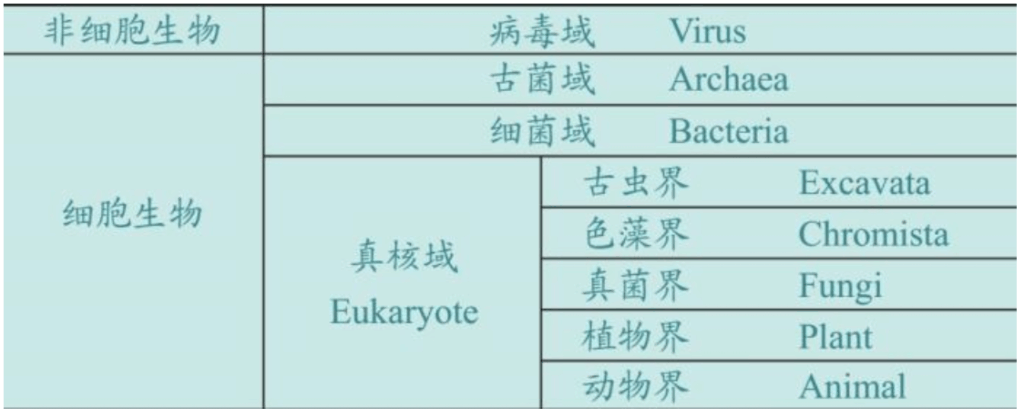
马古利斯和她儿子联合出版了24本论著,揭示的事实佐证了洛夫洛克的理论:35亿年前,地球出现第一批微生物厌氧菌,它们吃下二氧化碳,吐出“毒气”氧气,导致地球早期大气层出现戏剧性的变化。氧气的出现彻底改变了地球的物理——地质——化学——生物过程,地球与邻近的金星和火星变得完全不同了。这些厌氧菌的一部分后裔,演化为具有光合作用的有机组织——植物。25亿年前,地球出现第二批微生物,好氧菌们吃下氧气,吐出“毒气”二氧化碳。厌氧菌演化为植物,好氧菌演化为动物,分解植物和动物尸体的是菌类和细菌。
马利古斯告诉我们,所有生命来自一个起源,至今使用着同一套生命系统:DNA——RNA——蛋白质系统。所有生命都是互惠互利——互相协助——共同演化,最终形成覆盖在地球表面的一层生物圈。DNA的结构都是一样的:基因存贮在DNA里。
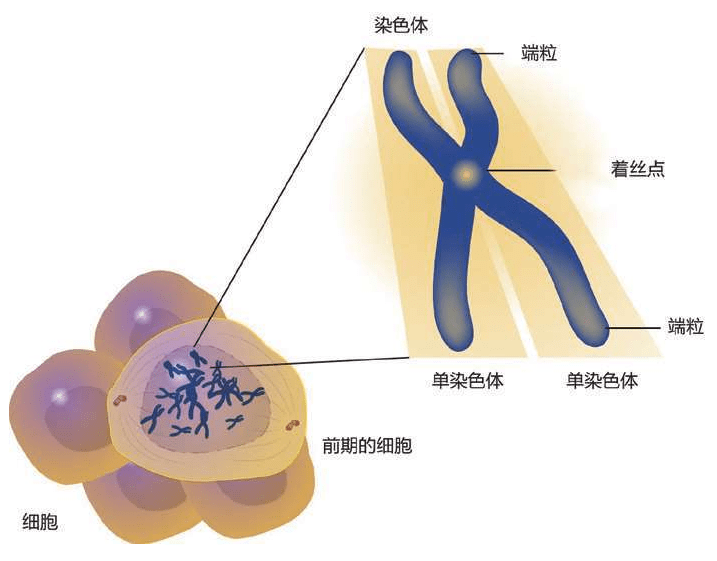
人类的DNA太长了,大约2米,很难存放和复制,于是被 “切割”成46个片段(其他生命的染色体数量不一),人类每个细胞的细胞核里都有一个染色体组(基因组)。过去认为人类约有100万亿个细胞,所以一个人的染色体DNA长度约为200万亿米,即2 000亿千米。地球到月球的距离38万千米,亦即一个人的DNA总长度可以在地球——月球之间来回53万次。现在的新研究认为,人的细胞数量大约为1亿亿个,这是原来估计数量的100倍,现在已经很难估算一个人的DNA的总长度能够在地球——月球之间来回多少次。
如此复杂的DNA存储器,可以存放非常复杂的遗传信息和生命程序——基因。在长达35亿年的时间里,地球生命的演化过程跌宕起伏,多次经历盛衰兴亡,除了5次著名的大灭绝之外,还有十几次小的灭绝事件。但是,DNA既未消亡,也未改变,只是基因(生命的程序及其传承)越来越复杂,多次合并共生使得地球的生命五彩缤纷,欣欣向荣。
马古利斯的生物演化假说得到了越来越多的证明,考古证据发现地球上出现的生命形式几乎难以计数,至少出现过300亿种生命物种,微生物的种类更是无法统计。很多科学家甚至开始设想,人类如何殖民到另外的星球?他们设想,首先将厌氧菌和好氧菌送到一个合适的星球,逐渐改变这个星球的大气层,形成极地冰盖——植物开始生长——云团出现——反照率改变,从而将这个没有生命的死星球从静止不变的状态转换变化成为一个美丽的、活的、呼吸的、演化的整体。
这就是盖亚的力量。
盖亚理论蕴含着富有诗意的丰富内涵,具有重大的指导意义。整个人类开始审视与大自然的关系,提出了保护环境、保护臭氧层、减少二氧化碳排放、防止气候变暖等一系列的新观念和新理论。
盖亚理论指出了生命的起源,生物科学和DNA的研究结果证明各种生命的起源和演化关系的的确确是息息相关的,人类(以及哺乳动物)的主要能量不是来自每日三餐,而是来自线粒体使用氧气制造的高能ATP能源……
如果人类违反大自然的规律,对抗DNA中蕴藏的基因,人类不仅会罹患更多的疾病,还会破坏地球的生命存在的条件,甚至再次引发生命的大灭绝。
结语
21世纪是生物世纪。
2000年6月26日宣布的“人类基因组工程”的首要目的是揭示各种疾病的秘密,找到治疗这些疾病的方法,以及查明500多种遗传病的起因……但是,人类对基因的了解刚刚起步,对大自然的精妙设计仍然一无所知。的确,我们非常聪明,研发出各种新设备仪器、新药品、新疗法……试图对大自然赐予的基因修修补补,甚至造出更好的下一代,但是我们必须考虑深远的长期后果。
仅仅两个世纪之前,我们作出开发煤炭、石油、天然气、油砂等化石能源的重大决定,后来发现石油制造的农药和化肥可以增加粮食产量……当时谁也没有预料到,这一决定导致了两个世纪后的人口爆炸、大气变暖、环境恶化等一系列全球性恶果。
人类与基因组之间存在的最大鸿沟在于:我们完全不知道基因组如何传递进入生物体系;不知道各个基因如何正确地打开和关闭,从而在细胞中进行生物化学物的混合和制造;不知道这些细胞如何懂得构建出一个生理组织,这些生理组织又如何自我协调而成为一个切实有效的有机系统?这个系统如此复杂,在这些不可理喻的总体联合作业面前,人类除了震惊,已经无法理解。
我们以有限的可怜的遗传基因的修补手艺,试图修修补补的最可能结果,将是失败大于胜算,亏损大于收益,风险无法估量。我们最安全的出路是弥合我们与大自然决裂,解决进入贪婪无度的农业社会至今带来的一系列社会问题。
2012年9月5日,人们又一次发现自己错了。
2012年9月5日开始,世界第一大媒体《时代》的一篇报道的题目本身就蕴含认错的含义:《垃圾基因:其实并非无用》(Junk DNA-Not So Useless After All)。这篇报道连续5天占据《时代》网络版头版位置。
这个消息,也是世界所有媒体的头版新闻。
2012年9月5日,总计30篇论文同步发表于《自然》(Nature)、《科学》(Science)、《基因研究》(Genome Research)等杂志上。
这是ENCODE第一次公布研发成果,并且是全球同步公告。
ENCODE是“DNA元素百科全书工程”(Encyclopedia of DNA Elements,ENCODE)的简称,是从2003年开始,全世界32个研究机构联合进行的一项巨大工程,目标同样是人类基因组。
2003年,人类基因组测序完成。编码的遗传基因只有2.1万——2.3万个,在全部约30亿个碱基对中所占的比例不足2%。它们是制造人体的全部蓝图和组装手册,包括人体的各种蛋白到每一个生物组织、器官、五官、皮肤、外观等的制造和装备。而其余的98%以上的基因不参与编码,即不涉及人的制造。所以,2003年基因组工程完成时,很多人感到相当失望,“垃圾基因”流行一时。与此同时,很多人也不相信大自然会犯下如此的错误。
事实上,这里依然是人类知识的荒漠——我们对基因的了解才刚刚开始。
10年后,2012年9月5日,30篇同步发布的论文告诉大家,在占整个基因组碱基对的98%以上的非编码基因中,80%以上具备生物化学活性,它们不是无用的垃圾,它们具备人类过去无法想象的功能。这80%DNA中包括400多万个基因开关(gene switches),每个细胞都能通信联络。这80%DNA中隐藏的指令,虽然不参与人体组织和蛋白的制造,但是却控制着我们的人生,例如大脑神经元的生长、吃肉以后指令胰腺分泌胰岛素、下令某些皮肤细胞死亡的同时产生新的皮肤细胞……这些DNA指令就像一场不可思议的人生舞台设计的全套剧本,具体到每一个演员(细胞)的出场和每一个舞蹈动作。参加这场人生演出的“演员”,过去只有DNA、RNA和蛋白,现在,各种新陈代谢因子和精巧绝妙的设计也参与了进来。科学家们已经无法用语言向纷至沓来的记者描绘这些新发现……
“DNA元素百科全书工程”不仅打开了疾病治疗的新大门,也对人类进化有了更多了解——我们的头发与脚趾甲,到底是怎么生长的?我们的基因组,难道一直在操作着这些生长?现在科学家可以了解“何时”与“何处”的基因开关表达,直到整个人体的全部功能,这是错综复杂到不可思议的一场人生舞台表演。
人类基因组工程之后,科学家已经从基因组里找到心脏病、糖尿病、精神分裂、孤独症等疾病的线索,但是还有几百种疾病的线索找不出来。现在,科学家终于可以在更加扩大的非编码基因范围里寻找新的线索。科学家们现在已经可以说,他们找到了以前无法完全理解的基因影响疾病的更多线索。
人类曾经犯过很多错误,但是最后都发现了真相,纠正了错误。
17世纪,伽利略提出“日心说”推翻了教会权威努力维持的地心说,证明我们不是宇宙的中心,告诉我们必须以开放的心态对待那些反对“历史事实”或“历史实践”的思想。
英国和法国建立科学社会的办法是建立各种论坛,自由讨论科学方法——自然界所遵循的各种规则,构成了大自然的法则,这些大自然的法则是可以被发现,也可以被检验的。牛顿是把这些法则整理成为一个体系的第一个人。
20世纪唯一可以与牛顿相比的是爱因斯坦,他在1905年发表的相对论否定了绝对时间的存在。起初人们认为相对论太过抽象,没有实际用途。但是根据爱因斯坦相对论中质量与能源相互转换的理论,人们制造出了原子弹。
伽利略认为我们不是宇宙的中心。
牛顿是把自然法则整理为体系的第一人。
爱因斯坦否定了绝对时间的存在。
科学技术的发展历史一再证明,人类必须向新的理论开放。
幸存的物种,
不是最强大的,
也不是最聪明的,
而是最能适应变化的。
——达尔文(达尔文从来没有使用过“进化”一词,他认为进化意味着进步)
从希腊时代的早期起,哲学家的工作就是提出问题和思考问题,现代公认的最伟大的两个提问者是弗洛伊德和达尔文。
弗洛伊德(1856-1939)是第一位科学地研究潜意识的人。通过生理学的方法,他研究了病人自由思想时所表达的含义,得出结论:人的行为直接联系其童年时代的经验或被压抑的性幻想的潜意识记忆。这个结论震撼了世界,过了仅仅几十年,就成为现代文化的内核,也解释了人类如何从2 000人左右的小小群体成为地球的霸主。
达尔文的理论,经过一百多年的反复验证后才被接受:所有生存的和消亡的物种都是自然选择的结果。在认识达尔文理论的过程中,人类犯了无数错误。
这些历史给整个人类的最大的教训就是:我们必须向所有新的理论开放,因为新的理论可能被新的事实证明。
附录
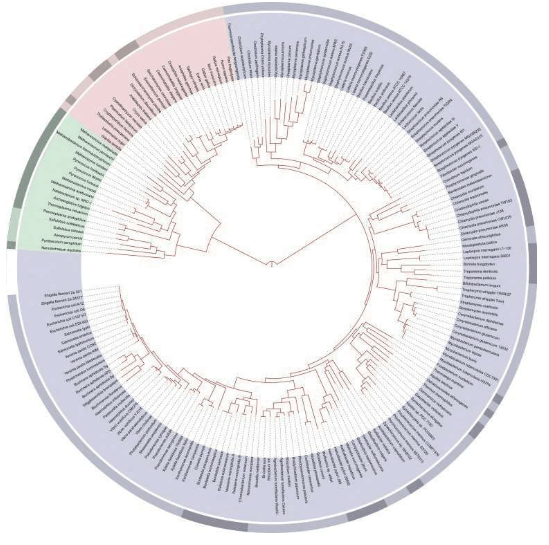
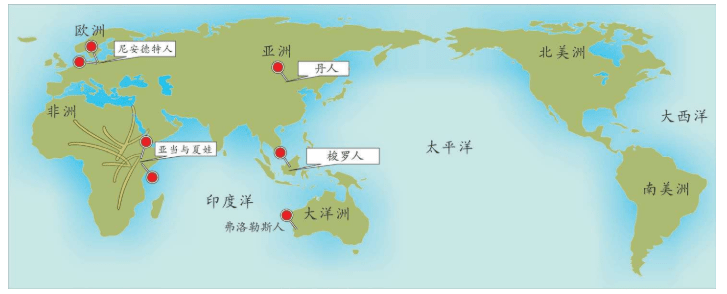
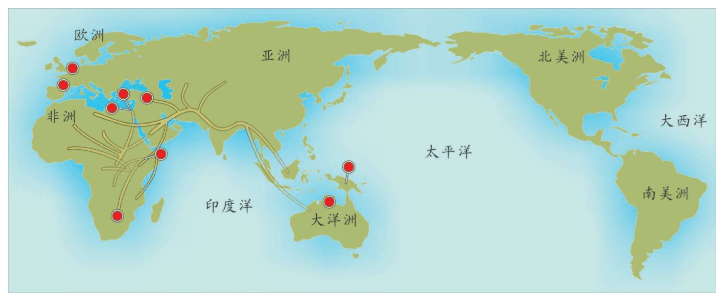
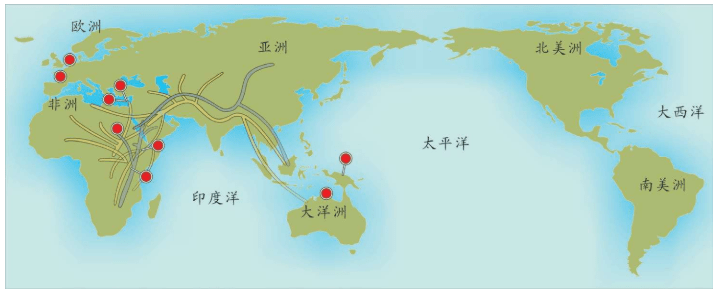
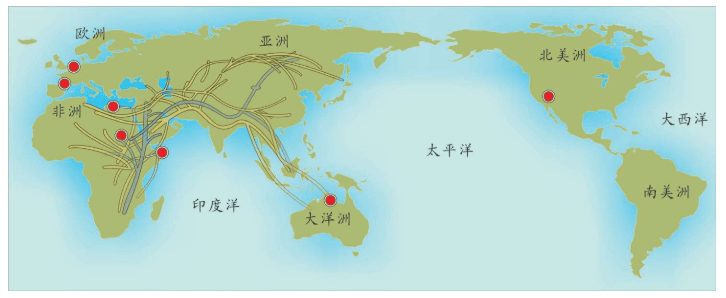
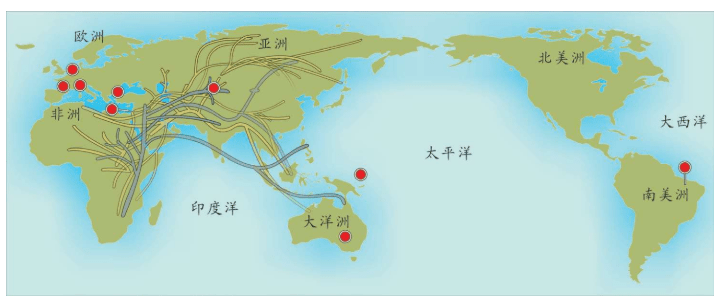
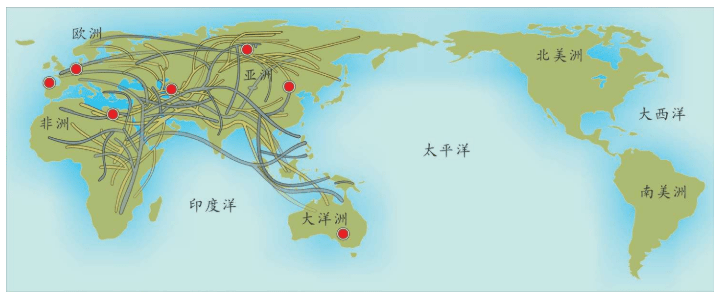
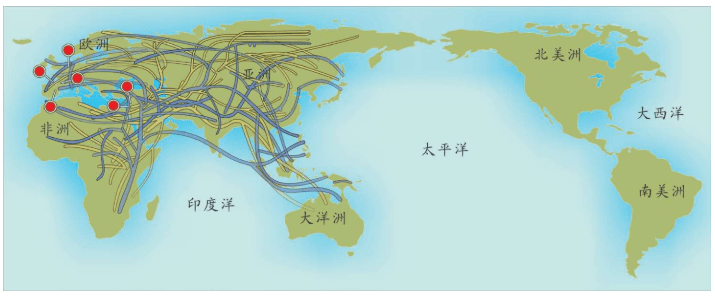
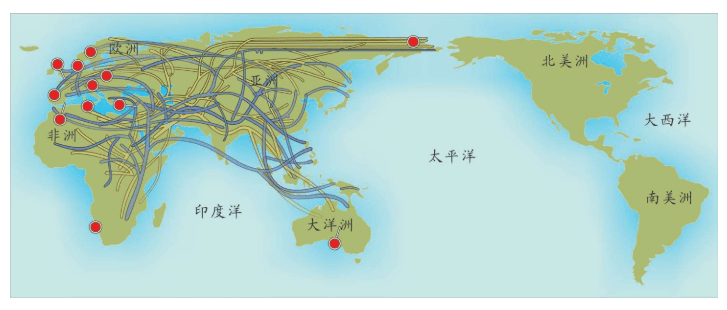
附录1 人类20万年的旅程图
现在的各种人类旅程路线,正在越来越详细。人类找出自己的来源,找出先祖,找到自己的根是一个长期的理想。而这个理想,随着越来越先进的测序设备和软件的发展正在逐步实现。经过大量采样和多次重复DNA测序,可以找到每一个体的DNA序列,从而推断这个个体所属的单倍群。
基因图谱工程采集的世界样本数量世界第一,所以绘制出的人类20万年以来的旅程路线最有权威性。这里列出11张美国国家地理网站公布的人类旅程图,能够更加详尽地解释人类走出非洲之后的路线。
附录2 主要单倍群简介
全世界的人类都属于一个单倍群,一个先祖氏族。这个氏族的单倍群遗传标记使我们可以研究迁移到世界各地的人类。人类群体可以通过遗传形态的计算分析出不同的单倍群。下面列举主要单倍群及其早期旅程,包括线粒体DNA与Y染色体单倍群。
附录2.1
线粒体mtDNA单倍群
线粒体DNA的全球迁移图,最常见的单倍群分布如下:
非洲:L=L1,L2,L3
近东:J,N(N包括在A,B,F,H,I,J,K,P,R,S,T,U,V,W,X)
欧洲:J,K,H,V,T,U,X
亚洲:A,B,C,D,E,F,G,M(M包括在C,D,E,G,Q,Z)
美洲:A,B,C,D和少量的X
单倍群L1/L0
先祖血统:“夏娃”→ L1/LO
考古学和化石证据认为人类大约20万年前起源于非洲,但是直到5万——7万年前才显示出现代智人的特征。
线粒体夏娃是人类谱系树的女性先祖的根。夏娃的早期的后裔,在非洲之内的各地活动,最后分为两个血统,分别携带不同的突变。比较古老的一个群体定义为L0,线粒体树最早的一个分支。现代的所有人类都属于这个L0群体的不同的关联分支,亦即L0是最古老的一个血统。更为重要的是,现代遗传数据研究证明,属于L0的原住民全部存在于非洲地区,这个证据进一步支持人类起源于非洲的结论。(几乎所有2万年以前的现代人遗骸都出土于埃塞俄比亚及其周围,只有极少几个出土于非洲南部。埃塞俄比亚及其周围也是其他人科生物化石出土最多的地区)
此后,线粒体夏娃的后裔又构成另一个群体L1,与较早出现的L0同时存在于非洲。L1的部分后裔最终离开非洲,L0的后裔始终全部在非洲。
单倍群L0
可能在10万年前起源于东非一带。经过几万年的迁移,他们的早期先祖曾经在撒哈拉以南的非洲游荡。
现在,L0频率最高的群体是非洲中部的俾格米人(Pygmies)和非洲南部的科伊桑人(Khoisan,常称桑人)。
L0的频率,在非洲中部——东部——东南部为20%——25%,在非洲北部——西部较低。
2 000-3 000年前的班图人大迁移过程中,铁器从非洲西部传播到非洲其他地区,土生土长的L0被同化或取代了,这使得L1的后裔在非洲中部、东部的频率显得比较高。在大西洋奴隶贸易时代,很多古老的血统离开了非洲大陆,美国的很多L0的后裔的线粒体DNA与莫桑比克人相同。
总体上,北美洲和中美洲的频率分布与非洲的西部和中西部惊人地接近,证明他们起源于非洲。
南美洲的L0的频率,接近非洲中西部——东南部的频率。
L1的频率,在非洲西部较高,但是L1非常分散,在中部——东部也比较高,在某些阿拉伯群体里也可以找到,只是比例较低,包括巴勒斯坦人、约旦人、叙利亚人、伊拉克人和贝都因人。
单倍群L2
先祖血统:“夏娃”→ L1/LO→L2
L2个体存在于撒哈拉以南的非洲,与他们的祖先L0/L1一样,他们也存在于非洲中部到遥远的南部。L0/L1在非洲东部——南部占主导地位,L2的先祖则向其他地区迁移。L2个体是非洲频率最高,分布最广泛的mtDNA单倍群,分为4个独特的子单倍群:L2a、 L2b、L2c、L2d。其中L2a频率最高的地区是非洲东南部,L2b、L2c、L2d主要分布在非洲西部和中西部。L2d是最古老的,L2b和L2c后来多次分支。
估计L2群体起源于7万年前的一个母系先祖,最早出现在非洲西部——中西部,其分布非常广泛,使得判断L2的准确起源地区非常困难。L2被认为是班图单倍群的起源,东南非洲的班图人中,大约一半的遗传血统来自L2。2 000-3 000年前的班图族大迁移,使得L2(L1的后裔)在非洲的中部和东部的频率非常高。由于L2频率最高的西非地区是奴隶贸易的主要地区,L2成为非洲裔美国人的最主要血统,频率约为20%。
单倍群L3
先祖血统:“夏娃”→ L1/LO→L2→L3
L3的最近的共同先祖生活在大约8万年前,L3的人群在非洲处处可见。他们是最早走出非洲的现代人,原因可能是气候的影响。大约5万年前,欧洲北部的冰原开始消融,非洲的气候开始变得温暖和潮湿,撒哈拉的部分地区变得适于居住。各种动物开始向北方迁移,L3跟随着好气候和猎物也向北迁移,具体路线不详。
L3在北非的频率很高,在整个非洲的班图人里也都可以找到L3。一些L3向西迁移到大西洋沿岸,包括佛得角群岛(Cabo Verde)。一些L3继续向北,最后完全离开非洲大陆,他们的后裔在中东人群中占10%,其中两个分支的单倍群走向了世界其他地区。L3是非洲裔美国人的重要单倍群,在美国人中可以找到源于非洲西部的L3的大部分血统,而源于非洲中西部——东南部的L3的频率较低。
单倍群M
先祖血统:“夏娃”→ L1/LO→L2→L3→M
L3的后裔M单倍群离开了非洲,可能通过红海——亚丁湾(Gulf of Aden)一带渡过狭窄的海峡,从东非海岸来到阿拉伯半岛。这是一场长距离大迁徙的开始,M沿着中东——欧亚大陆南部——澳大利亚,最后到达波利尼西亚。M是出现在约6万年前的第一批走出非洲的人类。
M属于亚洲血统,在阿拉伯半岛东部频率很高,在阿拉伯半岛南部约15%,在Levant(地中海东部)地区不存在,在巴基斯坦南部和印度北部的频率高达30%——50%,在印度河谷以东呈现广泛的分布和更大的遗传多样性,说明携带M的人群是南亚第一批居民的后裔。
M有多个分支:M1是非洲分支,M2-M6在印度,M7在东南亚南部,M7的两个分支M7a和M7b2分别在日本和韩国。M7在中国南部和日本的频率约15%,在蒙古的频率较低。
单倍群M1
先祖血统:“夏娃”→ L1/LO→L2→L3→M→M1
携带M突变的人群离开非洲走向印度次大陆和亚洲东部时,M1没有向东走,而是返回了非洲。M1包括4个独特的突变,年代都在6万年左右,在东非形成4个分支,最近1万——2万年分离了。
现在,非洲东部的线粒体血统的20%属于M1,分布横跨红海两岸,占地中海的M血统的大部分,在尼罗河的所有血统中占7%。
M1在印度和亚洲东部很少见,有趣的是,印度和亚洲东部的M和非洲东部的M1的年代是相似的,所以估计M1是走出非洲后又返回非洲的人群。
单倍群C
先祖血统:“夏娃”→L1/LO→L2→L3→M→C
M分支出来的一批人群进入中亚广袤的干草原:约五万年前,C的第一批成员北上到达西伯利亚,他们中的一些人后来最终进入北美洲和南美洲。
C起源于里海和贝加尔湖之间的中亚大平原,属于西伯利亚血统,占西伯利亚地区的20%。由于年代久远,在欧亚大陆北部频率较高,被认为是最早定居在这一带的第一批人类。
C的后裔向四周扩散并迅速南下,进入亚洲的北部和中部,但是频率逐步下降,在中亚为5%——10%,在东亚约3%。向西迁移的C的后裔终止于乌拉尔山脉和伏尔加河,仅为1%,说明早期人类受地理因素影响很大。
在1.5万——2万年前,适应西伯利亚寒冷气候的C的后裔,跨过白令海峡来到阿拉斯加,现在占北美洲和南美洲的土著的20%。但是,既不清楚他们的具体迁移路线,也不清楚他们迁移了多少批次。
单倍群D
先祖血统:“夏娃”→ L1/LO→L2→L3→M→D
约5万年前,从M分支的另一批人群D单倍群进入中亚干草原——亚洲东部,他们的第一批成员继续向东,最终进入北美洲和南美洲。
与C单倍群一样,D单倍群也居住在里海和贝加尔湖之间的中亚大平原,属于欧亚大陆东部血统,D的后裔向四周扩散并迅速南下,现在是亚洲东部的重要血统,约占20%。D的频率向西的方向逐步减少,在亚洲中部为15%——20%。
D是北美洲和南美洲土著的5类线粒体DNA之一。
单倍群Z
先祖血统:“夏娃”→ L1/LO→L2→L3→M→Z
约3万年前,Z单倍群的第一个成员北上进入西伯利亚,开始向亚洲东部的旅程。Z属于西伯利亚血统,居住在里海和贝加尔湖之间,现在约占这一地区的3%。
Z单倍群向四周扩散并南下进入亚洲的北部和中部,现在约占亚洲东部的2%。但是,Z向其他方向的迁移似乎都失败了。
当C和D的后裔进入美洲时,Z的后裔没有前往美洲。由于C-D-Z的居住区域相同,也许Z也进入了美洲,但是这支血统在美洲绝嗣了。
单倍群N
先祖血统:“夏娃”→ L1/LO→L2→L3→N
N与M是L3的两个直接后裔分支。M来自第一波走出非洲的大迁移,N来自第二波走出非洲的大迁移。M的路线是通过红海,N的路线是沿着尼罗河,通过西奈半岛走出非洲,因为沿着尼罗河谷地迁移可以找到足够的食物与饮水。这些L3的后裔最终构成了N单倍群。
N的早期成员离开撒哈拉沙漠的严酷环境,生活地中海东部和亚洲西部,可能当地依然存在着尼安德特人,因为以色列卡巴拉洞穴(Kebara Cave)出土了约6万年的尼安德特人遗骸,说明两种人科生物当时都在地中海沿岸。
携带N单倍群的突变特征的一些成员组成很多群体,向亚洲——欧洲——印度——美洲等地进发。N被认为是欧亚大陆西部的单倍群,因为在中东和欧洲的几乎所有线粒体血统中都发现了N。
单倍群N1
先祖血统:“夏娃”→ L1/LO→L2→L3→N→N1
N单倍群的地理分布广泛,其后裔N1是德裔的4个犹太血统(Ashkenazi)之一。公元1300年,德裔犹太人约2.5万人,20世纪达到850万人。在非德裔犹太人中,很少见到N1。现在N1的德裔犹太人约80万人,为4个德裔犹太人线粒体中第二大血统。N1也出现在Levant——中亚——埃及人群中。
单倍群A
先祖血统:“夏娃”→ L1/LO→L2→L3→N→A
约5万年前,A单倍群的第一个成员越过西伯利亚,最终来到北美洲和南美洲。A可能起源于中亚高原,然后扩散到亚洲东部几个地区。在美洲土著中第一次发现A,使得遗传学家开始用这个突变研究史前人类的迁移。
除了极少的例外,A是爱斯基摩人的唯一血统,A也是西伯利亚——阿拉斯加——加拿大的美洲土著的血统,可靠的起源时间约为1.1万年前。这个时间成为一个分子钟,用于估算爱斯基摩人和美洲土著的迁移时间,但是不能用于估算世界其他地方的人类迁移时间。
单倍群B
先祖血统:“夏娃”→ L1/LO→L2→L3→N→B
约5万年前,B单倍群的第一个成员进入亚洲东部,最终来到北美洲和南美洲,以及波利尼西亚的大部分地区。这个B单倍群可能起源于里海和贝加尔湖之间的中亚高原,成为亚洲东部的创始血统之一,B、F、M构成了现在亚洲东部所有线粒体血统的大约四分之三。
B单倍群向四周扩散并迅速南下进入亚洲东部,现在约占东南亚的17%,约占中国全部基因池的20%,并且广泛分布在太平洋沿岸,从越南到日本,少量存在于西伯利亚土著(约3%)。由于历史久远,频率较高,B被广泛承认是欧亚大陆最早的人群之一,也是南北美洲的5个线粒体血统之一。
B单倍群的分支之一B4,从东南亚向波利尼西亚扩张。B4积累了在欧亚大陆的突变,最近不到5,000年内扩散到波利尼西亚,其中的一些中间血统出现在越南——马来西亚——婆罗洲,支持B4起源于东南亚的可能性。
单倍群I
先祖血统:“夏娃”→ L1/LO→L2→L3→N→I
I是N的后裔,起源于中东地区,在欧亚大陆北部和北欧地区呈现显著的多样性,所以I的早期成员在旧石器时代的中期可能已经第一次进入欧洲。
这一波进入西欧的移民潮称为奥里尼雅克文化(Aurignacian culture),他们的工具先进、首饰精美、社会组织也比较复杂。
他们留下的欧洲血统,在旧石器时代约为10%,在新石器时代约为20%,其余的欧洲血统(包括I)是在旧石器时代中期的2.5万年前进入欧洲的,在1.5万年前的冰河时代后期开始扩散。
单倍群W
先祖血统:“夏娃”→ L1/LO→L2→L3→N→W
W是N的后裔,从中东进入欧洲。与I类似,现在中东的W后裔比北欧的W后裔呈现更显著的多样性,说明W在中东居住更久,积累了更多突变。
也与I类似,W的后裔是在旧石器时代中期进入欧洲的,他们也参与创造了奥里尼雅克文化(Aurignacian culture)。
单倍群X
先祖血统:“夏娃”→ L1/LO→L2→L3→N→X
X主要有X1和X2两个分支,分布广泛且没有规则。
X1大部分位于非洲的北部和东部。X2广泛分布于欧亚大陆西部,在欧洲占2%,在近东——高加索——地中海地区比例升高,有的欧洲地区达到10%——25%,扩散时间约为1.5万年前。
X也是美洲土著的5个线粒体血统之一,但是仅存在于北美洲。5个美洲线粒体血统A、B、C、D、X中,唯有X没有完全出现在亚洲东部,原因不详。
单倍群R
先祖血统:“夏娃”→ L1/LO→L2→L3→N→R
R是N的一个后裔,这个女人又被称为欧亚大陆西部血统的共同先祖,R的频率较高的地区在伊朗——高加索——安纳托利亚(Anatolian,又称小亚细亚,位于土耳其)地区。
R的历史复杂,起源很早,分布很广泛,属于第二波走出非洲的人类。从中东地区开始,R与她的先祖N一起走过了上万年,凡是发现N的地点和时间几乎都同时发现R。两者的故事很难分辨。
R的一部分后裔走到亚洲中部——印度河谷,R的另一部分后裔在3.5万年前进入欧洲,成为第一批抵达欧洲的克罗马农人,当时尼安德特人仍然生活在欧洲。R的后裔现在是欧洲最重要的线粒体血统,超过75%。
单倍群F
先祖血统:“夏娃”→ L1/LO→L2→L3→N→R→F F起源于里海和贝加尔湖之间的中亚高原,属于亚洲东部创始血统之一。F、B、M构成了现在亚洲东部所有线粒体血统的大约四分之三。
F是R的后裔,起源于中亚和东南亚。大约5万年前,F的第一个成员进入亚洲东部,然后扩散到整个东南亚,现在占东南亚的25%以上。
F的多样性在越南最显著,F广泛分布于菲律宾——中国台湾土著——东南亚等太平洋沿岸,最北方延伸到西伯利亚中部的鄂温克人(Evenks),最南边延伸到婆罗洲的卡达赞人(Kadazan people)。在巴布亚新几内亚的某些沿海群体里也发现了F,这个单倍体可能也影响了印度尼西亚人的起源。
由于F在美拉尼西亚(Melanesia)——波利尼西亚地区的频率很低,不太可能是澳大利亚土著带给他们的,估计是6 000-8 000年前的汉藏语系群体的扩张期间,通过东南亚带到了美拉尼西亚——波利尼西亚地区。
现在,东南亚——印度尼西亚——美拉尼西亚——波利尼西亚等地的史前历史,还有很多有趣的谜团,有待遗传学家和考古学家解开。
单倍群pre-HV
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→pre-HV
pre-HV在红海周围处处可见,广泛分布在近东地区,属于埃塞俄比亚——索马里的共同起源血统,在阿拉伯国家的频率最高。这些pre-HV接近欧亚大陆西部,很多群体生活在非洲东部,这些人可能是后来返回了非洲大陆。与此类似,N和R的后裔也有一些人返回了非洲大陆。
pre-HV是R的后裔,有时命名为R0,在安纳托利亚——高加索——伊朗地区的频率也很高,在印度——巴基斯坦边境的印度河谷地区也有pre-HV,可能是近东的群体向东方迁移的结果。
还有一些携带pre-HV的群体进入欧洲成为克罗马农人,形成两个非常重要的欧洲线粒体血统:H和V,时间约为2万年前。这就是名称pre-HV的来源。当时他们人数很少,不断扩大的冰原把他们挤压到西班牙南部、意大利和巴尔干半岛。1.2万前气候开始变暖之后,他们开始向欧洲北方扩张。
单倍群HV
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→pre-HV→HV
HV是pre-HV的后裔,一组独特的突变定义了HV单倍群。虽然一些后裔血统前往中亚——印度河谷等地,或返回非洲,HV的先祖始终留在近东。
约3万年前,HV的一些成员翻过高加索山脉,进入欧洲,1.5万——2万年前,他们被冰原挤压到伊比利亚半岛——意大利——巴尔干躲避严寒,人口急剧减少,原先在欧洲形成的多样性也丢失了。冰原撤退时,他们重新向欧洲西部殖民,其中两个最常见的线粒体血统是H和V,它们存在于75%以上的欧洲血统里。
单倍群HV1
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→pre-HV→HV1
HV1是pre-HV的后裔,形成于3万年前。与HV类似,HV1在近东的频率也很高,包括安纳托利亚(现土耳其)——高加索山区,主要集中在俄罗斯南部和格鲁吉亚。其中一些成员越过高加索山脉进入俄罗斯南部,来到黑海的干草原,然后向西进入波罗的海各国和欧亚大陆西部。今天,这些HV1的后裔血统位于东欧诸国和地中海东部地区。
虽然距离伊比利亚半岛不远,HV1的后裔与H和V的后裔没有发生关系。非常有趣的是,在非洲东部也发现了HV1的后裔,尤其是埃塞俄比亚,这很可能是最近2,000年来的奴隶贸易的结果。
单倍群H
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→pre-HV→HV-H
冰河期结束,人类再次向欧洲殖民时,出现频率最高的单倍群是H,H构成欧洲女性基因池的40%——60%。罗马和雅典的H的比例为40%,西欧其他地区的比例也差不多。越向东走,H的比例越低。土耳其的比例约25%,高加索山区的比例约20%。
H不仅是欧洲西部的主要单倍群,在东方也发现了H:东南亚约20%,亚洲中部约15%,亚洲北部约5%。更加重要的是,H在东方和西方的时间不同。在欧洲,H的时间估算为1万——1.5万年,实际上,H在3万年前已经进入欧洲,但是冰河时期人口急剧减少,多样性也丢失了,所以计算出来的时间比较短;在亚洲的中部和东部,H的时间估算约为3万年,亦即H血统很早以前也从近东迁移到亚洲。
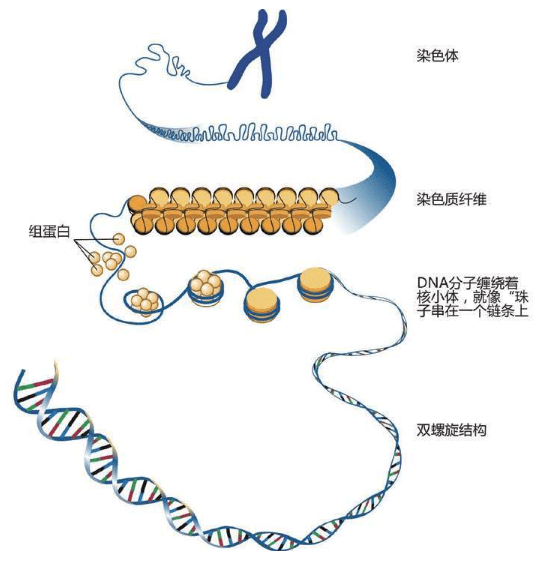
单倍群V
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→pre-HV→HV-V
现在,V局限于欧洲的西部——中部——北部,时间约1.5万年。这意味着人类在冰河时期,曾经在欧洲南部的避难地逗留了大约5 000年。
在西班牙北部相对封闭的巴斯克人(Basques)中,V的比例约12%。在其他西欧群体中,V的比例约5%。阿尔及尔和摩洛哥也发现了V,说明伊比利亚半岛的群体曾渡过直布罗陀海峡来到北非。有趣的是,斯堪的纳维亚(Scandinavia)北部的Skolt Sami人中V的比例最高。Skolt Sami人是狩猎采集群体,他们随着驯鹿,季节性地在西伯利亚和斯堪的纳维亚之间来回游牧。
单倍群J
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→J
J是R的后裔,这个女人出现在大约4万年前,这个单倍体是新石器时代人口大扩张中最重要的单倍群之一,她的后裔分布非常广泛:在印度——巴基斯坦、阿拉伯半岛、欧洲东部和北部都有J。
J在近东的多样性远远高于欧洲,J是黎巴嫩沿岸很多群体的先祖,在阿拉伯的比例最高:贝都因人和也门人中占25%
单倍群K
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→K
K也是R的后裔,这个女人出现在大约2万年前,她的后裔分为几个不同的分支,地理上极其分散,在欧洲、北非、印度、阿拉伯、高加索北部、斯堪的纳维亚、近东等地区都有。还有很多K的后裔进入俄罗斯南部的黑海干草原。
与N1类似,K也是一个著名的单倍群,因为K和K的分支涵盖了德国裔犹太人(Ashkenazi)的4个线粒体DNA单倍群中的3个,相当于300万个德国裔犹太人。
但是K在非德国裔犹太人中的频率比较低。在地中海沿岸Levant——中亚——埃及地区,K的比例约3%。
单倍群T
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→T
T也是R的后裔,这个女人出现在大约4万年前。T的分布广泛,最南边到阿拉伯半岛,最东边到印度河谷。T出现于旧石器时代,也是新石器时代大扩张的主要单倍群之一。
单倍群U
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→U
U也是R的后裔,这个女人出现在大约5万年前,T和T的分支广泛分布在欧洲、北非、印度、阿拉伯、高加索北部、近东等地。其中最重要的一批群体翻越高加索山脉,进入黑海干草原,然后继续西进,抵达现在的波罗的海各国和欧亚大陆西部。
U在欧洲常见,在地中海东部的频率约7%。
单倍群U5
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→U5
U5是U的后裔,起源于5万年前。局限于斯堪的纳维亚,尤其是芬兰。原因可能是芬兰的地理位置相对比较封闭。
季节性跟随驯鹿迁移的游牧群体Skolt Sami人中,U5的比例高达50%。
U5也出现在北非摩洛哥——塞尔加内——阿尔及利亚的帕帕尔人群体中。在距离如此遥远的两个地区发现同样的遗传血统,确实出乎预料,这可能是冰河期结束之后,一场延续了大约1.5万年的大迁徙的结果。
U5在近东很少,比例约2%,在阿拉伯地区找不到。U5还出现在土耳其人——库尔德人——亚美尼亚人——埃及人里,这可能是欧洲人回流到近东的结果。
单倍群U6
先祖血统:“夏娃”→
L1/LO→L2→L3→N→R→U6
U6是U的后裔,起源于5万年前。U滞留在近东,但是U的一部分后裔北上进入欧洲和斯堪的纳维亚,还有一部分后裔沿着地中海沿岸向西走,现在北非地区的U6约占10%。
U6的文化相当发达,石器和骨器先进,首饰和雕塑精美,岩画也很著名。冰河期结束之后,U6的先祖渡过直布罗陀海峡,进入西班牙和法国南部,他们来往于北非和欧洲南部,所以在两边都留下了血统。
附录2.2Y染色体单倍群
进入南北美洲的Y染色体单倍群只有两个:C3和Q3。C3属于第一波走出非洲的群体,Q3属于第二波走出非洲的群体。
单倍群A
先祖血统:“亚当”→ M91
单倍群A起源于约6万年前,由M91定义。遗传多样性随着时间增大,所以M91联系着一个更早的男性共同先祖“亚当”。
现在,埃塞俄比亚——苏丹——洲南部很多人携带着M91,他们的文化传统仍然关联着先祖的生活方式。例如,喀拉哈里沙漠(Kalahari)的桑人——布须曼人(San Bushmen)和坦桑尼亚哈扎人(Hadza),都继续使用嗒嘴音。
距今2 000-3 000年的非洲班图人的文化大扩张,大大挤压了A单倍群的人口数量和古老文化。
单倍群B
先祖血统:“亚当”→ M60
M60定义了B单倍群,一个起源于5万——6万年前的古老非洲血统。与其他古老的血统后裔类似,B的分布非常分散,遍布整个非洲大陆,涉及很多不同的人群和文化,包括俾格米人:Biake people人和Mbuti people人。
单倍群C
先祖血统:“亚当”→M168→M130
约5万年前,可能在南亚,一个男人出生时携带着基因标记M130,他的晚近先祖参与了第一批走出非洲的旅程。这批人沿着非洲海岸前进,路线大致为阿拉伯半岛南部——印度——斯里兰卡——东南亚。其中一些人渡海抵达并定居澳大利亚。这批早期移民沿着海岸线旅行,不需要什么新技术,依靠海洋资源维生,不到5 000年就迁移到澳大利亚。
在这部分迁移的人群中,并非每个人都前往了澳大利亚,许多人留在东南亚沿海,逐步向内陆迁移,几千年后进入亚洲东部,包括蒙古和西伯利亚。在大约1万年前,这批群体的一部分居住在中国北方或西伯利亚东南方的后裔,乘船沿着太平洋海岸线迁移到北美洲,证据就是北美洲的纳——德内语系(Na-Dene languages)。
这个语系仅限于北美洲的西半部。在纳——德内语系的群体中,尤其是加拿大西部和美国西南部的男性美洲土著中,携带基因标记M130的比例为25%。
单倍群YAP
先祖血统:“亚当”→M168→YAP
YAP是Y染色体Alu多态性(Y Alu Polymorphism)的简称,Alu是Y染色体上长度约300碱基对(核苷酸)的一个区段,又称阿鲁元素(Alu element),这个无害的Alu重复地插入人类基因组的不同部位,插入模式已经超过100万种并遗传给后裔。约5万年前,一个男人体内的Y染色体上出现了这个300碱基对的区段并遗传给他的后裔。
YAP在非洲东北部,属于撒哈拉南部3个最常见的遗传分支之一。YAP血统后来分支成为2个距离遥远的群体:D单倍群在亚洲,由M174突变定义;E单倍群主要在非洲和地中海地区,由基因标记M96定义。
单倍群D
先祖血统:“亚当”→M168→YAP→M174
D的先祖与C单倍群一起,构成离开非洲的第一批主要的移民潮。现在D存在于东南亚和安达曼群岛(Andaman Islands),但是印度没有。D的一部分后裔现在日本,还有一部分后裔现在西藏。
单倍群D1
先祖血统:“亚当”→M168→YAP→M174→Ml5
遗传标记M15定义了D1,这个单倍群最早出现于3万年前,地点可能在东南亚。这个血统的后裔后来进入西藏。现在D1存在于东南亚和西藏,西藏的频率最高。
单倍群D2
先祖血统:“亚当”→M168→YAP→M174→P37.1
约3万年前,第一次出现基因标记P37.1。这个标记定义了D2,这个M174的后裔单倍群逐步向北迁徙,最后抵达日本,现在D2是日本最常见的单倍群,在某些日本群体中的频率超过50%。
单倍群E
先祖血统:“亚当”→M168→YAP→M96
3万——4万年前,基因标记M96第一次出现在非洲东北,准确的地点尚不清楚。后来进入西非,并随班周大迁徙成为非洲主流。约5万年前,一个中东氏族开始走出非洲进行第二波大迁移,他们大部分是M89(参阅F)的后裔。这个中东氏族向北走,最后定居在中东。E单倍群也来到中东,具体路线可能与中东氏族相同,也可能不同。
单倍群E3a
先祖血统:“亚当”→M168→YAP→M96→M2
约3万年前,这个男人出现在非洲,他的后裔向撒哈拉以南迁徙。在大约2 500年前的班图人大扩张期间,E3a的后裔从非洲中西部向非洲的东部和南部扩散。现在,E3a在尼日利亚和喀麦隆的频率超过70%。在非洲裔美国人中,E3a也是最常见的血统。
单倍群E3B
先祖血统:“亚当”→M168→YAP→M96→M35
约2万年前,M35出现在中东。冰河期结束,气候变暖之后,人类从游牧的狩猎采集生活方式向定居农业生活方式转变。约8 000年前,新月沃土地区出现了成功的农业,向地中海周围扩张,社会组织也开始复杂化。M35单倍群可能以30-50人的小群体进行扩散。
单倍群F
先祖血统:“亚当”→M168→M89
约4.5万年前出现在非洲东北部或者中东的基因标记M89,现在存在于世界的90%以上“非非洲人”的身上。
最早离开非洲的人类沿着海岸线来到澳大利亚,但是F的路线是沿着大草原迁移,路线为非洲东部——中东——继续向前。F属于走出非洲的第二波移民,很多M89的后裔留在中东,还有很多M89的后裔追随着猎物的迁徙继续前进,途经今天的伊朗地区进入欧亚大陆中部广袤的干草原。
当时,这片巨大的干草原像一条“超级高速公路”,从法国直达韩国。他们从亚洲中部分别向西方和东方扩散。
单倍群G
先祖血统:“亚当”→M168→M89→M201
G的后裔都携带基因标记M201,这个男人大约在30,000年前出生在中东的东方地区,可能是巴基斯坦或印度的喜马拉雅山脚下。在新石器时代的农业人口大扩张之前,G的后裔生活在印度河谷地区。农业人口来到之后,取代或灭绝了很多G的后裔,幸存的G的后裔学会了农耕技术。
G有3个关联的“兄弟”单倍群:H,I,J,他们的部分后裔随着农业继续扩散。
单倍群G2
先祖血统:“亚当”→M168→M89→P15
基因标记P15出现于约1万年前,定义了单倍群G2,出现于中东,G2的后裔很快扩散,经过现在的土耳其进入欧洲东南部。
约1.5万年前的冰河期巅峰时期,早先的欧洲移民被冰原挤压和封锁在欧洲南部的几处避难地,人口较少。冰河期结束后,G2的各个血统从中东向欧洲的北部和东部扩散,还有一些人进入欧亚大陆的西部。
单倍群H
先祖血统:“亚当”→M168→M89→M69
约4.5万年前,起源于中东的H单倍群的先祖沿着欧亚大陆的干草原上的“超级高速公路”移民,后来辗转进入印度。约3万年前,一个携带遗传标记M69的男人诞生了,M69定义了这个新的H单倍群血统。虽然M69是一个“印度基因标记”,但是这个男性先祖的出生地点可能在中亚的南部,他的后裔是最早定居印度的内陆地区的群体之一。
这个单倍群H并非抵达印度的第一批人类。在5万——6万年前,第一批人类从非洲沿着海岸线来到印度,有些人定居在印度的沿海,但是内陆地区的大部分人类属于H单倍群的成员。
单倍群H1
先祖血统:“亚当”→M168→M89→M69→M52
M52定义了单倍群H1,这是印度的主要血统之一,M52在大约2.5万年前第一次出现在印度,属于迁移到印度的第二波主要人类群体。
H1的先祖也起源于中东,现在某些印度地区的频率为25%,在伊朗和中亚的南部,也有频率不高的H1。
单倍群I
先祖血统:“亚当”→M168→M89→M170
I的先祖是中东氏族M89的一部分,他们向西北的巴尔干迁移,后来扩散到欧洲的中部,在2.1万——2.8万年前,这些群体在欧洲的西部创造了格拉维特文化(Gravettian culture)。格拉维特文化是指法国的格拉维特发现的一些新的技术和艺术,石器工具也与早期的奥里尼雅克文化(Aurignacian culture)有所不同。格拉维特文化的狩猎工具更加先进,并且出现了女性雕塑的形象、贝壳首饰,以及动物骨头建造的房屋。(非洲喀拉哈里沙漠(Kalahari)的桑人,属于最古老的A单倍群)
这个共同先祖M170出现在约2.5万年前,在最后一次冰河期,他的后裔被迫退避到欧洲南部的封闭的避难地——巴尔干半岛和伊比利亚半岛(现西班牙)。
冰河期结束后,I单倍群的后裔在重新殖民欧洲时期担任了重要角色。
单倍群I1a
先祖血统:“亚当”→M168→M89→M170→M253
约2万年前,这个群体像很多欧洲群体一样退避到南部的避难地,以躲避冰河期巅峰时期向南扩张的大片冰原。I1a单倍群的避难地在伊比利亚半岛,其中一个男性成员出现了基因标记M253。
约1.5万年前地球变暖,I1A向欧洲其他地区扩散,在欧洲西北地区非常普遍,频率最高的地区是斯堪的纳维亚西部,可能很多维京海盗是I1A的后裔。
维京海盗多次入侵英国,这有助于解释M253在英伦诸岛的出现。
单倍群I1b
先祖血统:“亚当”→M168→M89→M170→P37.2
P37.2定义了单倍群H1b,这个基因标记约15 000年前出现在巴尔干,现在当地依然非常普遍。
P37.2可以识别冰河期退避到欧洲南部避难地的群体。
P37.2在冰河期结束后开始向欧洲北部和东部扩散,现在欧洲中部和东部非常普遍。这个血统可能是凯尔特人(Celtic)在公元前的一千多年的扩张。
单倍群J
先祖血统:“亚当”→M168→M89→M304
约1.5万年前,J诞生于新月沃土,这个地区包括以色列——西岸——约旦——黎巴嫩——叙利亚——伊拉克。现在J的频率最高的地区包括中东——北非——埃塞俄比亚。
在欧洲,J原先仅仅分布在地中海附近,农业出现后人口激增,J开始扩散。
例如,现在J和他的分支J2合计占犹太人的30%。
单倍群J1
先祖血统:“亚当”→M168→M89→M304→M267
新石器革命之后,随着农业的成功,J1和其他J单倍群在中东兴起,部分J1的成员前往北非并获得成功,证据是现在这一带的J1频率最高。
携带M267基因标记的其他J1的成员,一部分留在中东,一部分北上进入欧洲西部,但是频率较低。
单倍群J2
先祖血统:“亚当”→M168→M89→M304→M172
M172定义了起源于M89的J单倍群的主要分支。J2现在分布在北非——中东——欧洲南部,在意大利的频率是20%,西班牙南部的频率是10%。
单倍群K
先祖血统:“亚当”→M168→M89→M9
约4万年前,基因标记M9出现,地点在伊朗或亚洲中南部,这是中东氏族M89分支的一个新血统,这个血统的后裔经过3万年,扩散和成为地球的大部分人口。
这个大型血统K,被称为欧亚氏族,扩散过程延续了几万年时间,沿着欧亚干草原的“超级高速公路”,追随着猎物四处扩散,直到被亚洲中南部的巨大的山脉系统——兴都库什山脉——天山山脉——喜马拉雅山脉阻拦。这三大山脉的交会地区是帕米尔高原,位于今天的塔吉克斯坦,K单倍群在这里分为两支:一部分人北上进入亚洲中部,一部分人南下进入巴基斯坦——印度次大陆。帕米尔高原导致了K的分离。
今天北半球大部分人群的起源都可以追溯到这个男人M9:亚洲东部和北美洲的几乎全部,欧洲的大部分,以及很多印度人群。
单倍群K2
先祖血统:“亚当”→M168→M89→M9→M70
并非所有的M9都走向帕米尔高原,还有一些M9回到近东的舒适环境,这些人群中诞生了M70,时间约3万年前。
M70定义了K2,这个血统扩散到地中海沿岸各地,包括北非沿岸和欧洲南部的地中海沿岸。
有些人认为,K2就是腓尼基人,这些航海家建立了很多地中海沿岸的繁华的贸易据点,M70的起源可能在黎巴嫩一带。
现在,M70在地中海沿岸处处可见,在中东和非洲东北地区的频率是15%,在西班牙——法国的南部也可以找到M70。
单倍群L
先祖血统:“亚当”→
M168→M89→M9→M20
欧亚氏族M9的后裔可能在印度或中东诞生了M20。这个M20在约3万年前进入印度,形成了L单倍群,所以L又称为印度氏族。
印度南部的M20的频率达到50%,他们都是L单倍群的成员,虽然他们不是最早进入印度的群体。
单倍群M
先祖血统:“亚当”→M168→M89→M9→M4
由于冰河期的严酷气候,M4的先祖退避到东南亚的沿海地区。第一个携带M4的男人可能出生于1万年前,这个基因标记主要出现在美拉尼西亚(Melanesia)——印度尼西亚,少量存在于密克罗尼西亚(Micronesia)。
M4是随着水稻技术在各个海岛上传播的,当时中国和中东的农业正在发生。大约在4 000年前,水稻技术传播到婆罗洲——苏门答腊地区。然后,这里的群体携带着大量的淡水和水稻,航行到太平洋上的各个岛屿并定居下来。
单倍群N
先祖血统:“亚当”→M168→M89→M9→LLY22G
经过东亚的欧亚氏族诞生了基因标记LLY22G,定义了N单倍群,时间约1万年前,地点可能在西辽河流域,他的后裔成为江藏语系的部分和乌拉尔语系(Uralic)的群体,现在分布在斯堪的纳维亚的南部和亚洲的北部。
乌拉尔语系的文化,呈现出多样性:欧洲北部大部分人曾是狩猎采集群体,匈牙利人早期是牧马人,俄罗斯北方的很多群体也属于N。瑞典——挪威——芬兰——俄罗斯北部的Sami人是追随驯鹿的游牧——打鱼群体,人数仅剩下约8.5万人。
单倍群O
先祖血统:“亚当”→M168→M89→M9→M175
约3.5万年前,一个携带M175的男人出生了,地点在亚洲东部或中部。这些人是M9的后裔,他们当时在伊朗高原狩猎。
在冰河期的巅峰时期,O单倍群的先祖们抵达中国和亚洲东部,由于高山屏障的阻拦,他们封闭了上万年。O单倍群被称为东亚氏族:现在的亚洲东部,这个O单倍群的频率为80%——90%。在亚洲西部和欧洲, O单倍群不存在。
单倍群O1a
先祖血统:“亚当”→
M168→M89→M9→M175→M119
约3万年前,在中国南部出现的M119定义了单倍群O1a。他们可能是长江下游水到农业的人群后裔。
O1a的成员扩散到东南亚各地,他们的很多至今仍在东南亚。还有一些携带M119的群体进行了长距离的迁移,最终抵达中国台湾,现在台湾的几个原住民群体中的M119的频率高达50%。
单倍群O2
先祖血统:“亚当”→M168→M89→M9→P31
约3万年前,第一个携带P31的男人诞生了。P31定义了单倍群O2。这个男人居住在亚洲的东部,或许在中国南部。这个男人的后裔,有的向南扩散到东南亚,向东扩散到韩国和日本。这个基因标记现在是马来西亚——泰国等东南亚国家最常见的标记。
单倍群O3
先祖血统:“亚当”→
M168→M89→M9→M175→M122
这个携带M122基因标记的男人可能出生在中国中部,他的后裔分布非常广泛,约占中国男人的50%,所以,他的后裔很可能与农业的传播关系密切。单倍群O3的成员很可能都是长江中游种水稻和桑干河——永定河流域种粟米的农民的后裔。
亚洲东部的水稻农业的发达引发了大规模的人口扩张,考古学证据证明水稻技术一直传播到日本——中国台湾——东南亚,基因证据认为传播水稻技术的群体都携带着M122。
单倍群P
先祖血统:“亚当”→M168→M89→M9→M45
约3.5万年前,一个男人携带M45出生在亚洲中部,他是M9的后裔。
M9曾经扩散到一片富饶的大草原,包括今天的哈萨克斯坦——乌兹别克斯坦——西伯利亚南部。在冰河期的巅峰时期,草原环境日益恶化,迫使他们追随着驯鹿向北方进发,最后学会用兽皮搭建帐篷,改进武器和狩猎技术等。
M45是大部分欧洲人和几乎全部美洲土著的共同先祖。
单倍群Q
先祖血统:“亚当”→
M168→M89→M9→M45→M242
1.5万——2万年前,一个携带M242的男人诞生在极度严寒的西伯利亚。
M242的后裔成为第一批探索美洲大陆的人类。当时西伯利亚的气温很低,人类设法穿过没有积雪的苔原,来到亚洲的最北端——西伯利亚东部。
当时世界的海平面比现在低100米左右,西伯利亚——阿拉斯加之间形成了白令陆桥,人类可以直接走到美洲。
Q单倍群踏上了美洲的具体的时间还有争议,有人认为在2万年前。但是遗传学分析的数据是约1.5万年前,这个数据与考古证据基本吻合。
Q单倍群的成员进入北美洲之后开始南下,目前还不清楚他们如何越过北美的冰原。一种估计是落基山脉中的一条通道,另一种估计是沿着无冰的海岸线。
单倍群Q3
先祖血统:“亚当”→
M168→M89→M9→M45→M242→M3
第一批人类进入美洲后,M3出现了,时间是1万——1.5万年前,地点是北美洲。这是美洲土著中分布最广泛的一个血统,包括全部南美洲人口以及大部分北美洲人口。
M3在西伯利亚没有发现,仅仅分布在美洲。携带M3的群体继续南下,在大约1 000年里,就迅速抵达南美洲的南端。
单倍群R
先祖血统:“亚当”→
M168→M89→M9→M45→M207
在亚洲中部逗留了相当长的一段时间以后,经受着严寒气候考验的人类具备了更先进的技术,开始转头向西,进入欧洲大陆
这个氏族里的一个男人的Y染色体上,出现了一个新的突变M207,他的后裔分为两支:一部分人进入欧洲,一部分人后来转向南方,进入印度次大陆。
分析计算证实,这次迁移发生在1万年内,很多细节尚不清楚。
单倍群R1
先祖血统:“亚当”→
M168→M89→M9→M45→M207→M173
第一批大规模定居欧洲的群体的后裔是R单倍群的成员,M173定义了R单倍群:携带M207的群体一起向西迁移,半途分开。
约3.5万年前,M173的后裔抵达欧洲,当时尼安德特人处于衰亡时期。这些更聪明的M173的后裔获得更多的资源,可能加速了尼安德特人的灭绝。
在冰河期巅峰时期,M173的后裔撤退到南部的避难地:西班牙——意大利——巴尔干。冰河期之后,他们再次北上。现在欧洲的M173的频率非常高。
单倍群R1a1
先祖血统:“亚当”→
M168→M89→M9→M45→M207→M173
→M17
1万——1.5万年前,一个携带M17的男人诞生了,地点在现在的乌克兰或者俄罗斯南部。他的后裔继续游牧,并且驯化了马。他们骑着马,更加便于迁移:从印度到冰岛,他们迁移的范围很广泛。
在捷克——西伯利亚——亚洲中部,M17的频率约40%,在印度约35%,在中东地区为5%——10%。在伊朗,M17在伊朗西部仅为5%——10%,在伊朗东部高达35%。这些分布使得语言学家猜测:骑马的M17后裔可能是印欧语系的传播者。
单倍群R1b
先祖血统:“亚当”→
M168→M89→M9→M45→M207→M173
→M343
约3万年前,已经处于欧洲的群体中出现了遗传标记M343,定义了单倍群R1b。他们是克罗马农人的后裔,这些群体最著名的遗产是在法国南部等地留下的洞穴壁画。在此之前,人类的艺术作品主要是贝壳、骨头、象牙等材料制作的首饰。
单倍群R2
先祖血统:“亚当”→
M168→M89→M9→M45→M207→M124
约2.5万年前,亚洲中南部诞生了一个携带M124的男人,他的后裔迁移到巴基斯坦和印度东部。这个基因标记现在存在于印度北部——巴基斯坦——亚洲的中南部地区,频率5%——10%。这个单倍群R2也属于第二批大规模进入印度的移民,但是距离5万——6万年前的第一批大规模移民时间久远。(绝大部分2万年以上的现代人化石出土于苏丹——埃塞俄比亚——肯尼亚等东非地区,只有极少几个现代人化石遗骸出土在埃塞俄比亚以南的地区)
在欧洲东部的吉普赛人中,也发现了单倍群R2的成员,估计也是起源于印度次大陆地区。目前还不清楚这些远古时代的远程移民的细节。